Abstracts
Damming rivers to generate hydroelectric energy causes a series of changes in the environment, including impacts on the ichthyofauna. Knowledge of the fish community immediately downstream from a hydroelectric dam can help to reduce the negative effects of dam operation, such as fish entrance into the draft tube during turbines maintenance. We evaluated the temporal variation in fish community composition and abundance, near Três Marias Hydroelectric Dam (TMD), and its relationship with abiotic variables. Samples (n = 18) were carried out using gill nets in the tailrace of the dam, over the course of a hydrological year, at six time points during a 24-hour period. Abiotic data were also sampled to assess the relationship with the biotic data. In the tailrace, Characiformes showed the highest richness, while Siluriformes were the most abundant. Of the 33 species sampled, six were migratory. The fish community was significantly different between the dry and wet seasons, with higher abundances during the wet season. In the wet season, the frequency of migratory species in the community rose from 0.7 to 18.3%, and Pimelodus maculatus accounted for 14.5% of the overall abundance. We observed that Characiformes captures occurred mainly in the morning, with peak activity between 5:00 and 9:00 h. Siluriformes showed greater nocturnal activity and were largely captured between 17:00 and 21:00 h. Synchronizing operational maneuvers that represent high risk to the fish with periods of low abundance in the tailrace, i.e., during months of low precipitation, may be a measure to reduce the impact of the operation on the downstream fish community.
Fish kill; Ichthyofauna; Impact; Nictemeral; Operation
O barramento de rios para implantação de hidrelétricas causa uma série de modificações no ambiente, gerando impactos sobre a ictiofauna. O conhecimento sobre a comunidade de peixes encontrada imediatamente à jusante pode auxiliar na redução de impactos gerados pela sua operação. Assim, avaliamos as variações temporais na comunidade de peixes próxima à Usina Hidrelétrica de Três Marias quanto à composição de espécies, abundância e suas relações com variáveis abióticas. Foram realizadas 18 coletas com redes de emalhar no canal de fuga da usina, durante um ano hidrológico, em seis horários ao longo de 24 h, quando também foram obtidos dados abióticos. Foi observado que no canal de fuga da usina, Characiformes apresentou uma maior riqueza, enquanto os Siluriformes foram os mais abundantes. Das 33 espécies coletadas seis eram migradoras. A comunidade de peixes foi significativamente diferente entre os meses de seca e chuva, com maior abundância no período de chuva. Neste período, a frequência das espécies migradoras na comunidade subiu de 0,7 para 18,3%, sendo que Pimelodus maculatus representou 14,5% da abundância. A maior captura de Characiformes se deu no período da manhã com pico de atividade de 5:00 às 9:00 h. Os Siluriformes apresentaram maior atividade à noite, principalmente das 17:00 às 21:00 h. Conciliar manobras que oferecem mais riscos para os peixes com o período de menor abundância no canal de fuga, ou seja, nos meses de baixa pluviosidade, pode ser considerada medida para reduzir o impacto da operação sobre a comunidade à jusante.
Temporal variation in fish community in the tailrace at Três Marias Hydroelectric Dam, São Francisco River, Brazil
Raquel Coelho LouresI,II; Paulo Santos PompeuII
ICemig Geração e Transmissão, Gerência de Estudos e Manejo da Ictiofauna e Programas Especiais, Programa Peixe Vivo. Av. Barbacena 1200, 30190-131 Belo Horizonte, MG, Brazil. raquel.fontes@cemig.com.br
IIUniversidade Federal de Lavras, Campus Universitário, Departamento de Biologia /Setor de Ecologia, Pós-Graduação em Ecologia Aplicada, Caixa Postal-3037, 37200-000 Lavras, MG, Brazil. pompeu@dbi.ufla.br
ABSTRACT
Damming rivers to generate hydroelectric energy causes a series of changes in the environment, including impacts on the ichthyofauna. Knowledge of the fish community immediately downstream from a hydroelectric dam can help to reduce the negative effects of dam operation, such as fish entrance into the draft tube during turbines maintenance. We evaluated the temporal variation in fish community composition and abundance, near Três Marias Hydroelectric Dam (TMD), and its relationship with abiotic variables. Samples (n = 18) were carried out using gill nets in the tailrace of the dam, over the course of a hydrological year, at six time points during a 24-hour period. Abiotic data were also sampled to assess the relationship with the biotic data. In the tailrace, Characiformes showed the highest richness, while Siluriformes were the most abundant. Of the 33 species sampled, six were migratory. The fish community was significantly different between the dry and wet seasons, with higher abundances during the wet season. In the wet season, the frequency of migratory species in the community rose from 0.7 to 18.3%, and Pimelodus maculatus accounted for 14.5% of the overall abundance. We observed that Characiformes captures occurred mainly in the morning, with peak activity between 5:00 and 9:00 h. Siluriformes showed greater nocturnal activity and were largely captured between 17:00 and 21:00 h. Synchronizing operational maneuvers that represent high risk to the fish with periods of low abundance in the tailrace, i.e., during months of low precipitation, may be a measure to reduce the impact of the operation on the downstream fish community.
Key words: Fish kill, Ichthyofauna, Impact, Nictemeral, Operation.
RESUMO
O barramento de rios para implantação de hidrelétricas causa uma série de modificações no ambiente, gerando impactos sobre a ictiofauna. O conhecimento sobre a comunidade de peixes encontrada imediatamente à jusante pode auxiliar na redução de impactos gerados pela sua operação. Assim, avaliamos as variações temporais na comunidade de peixes próxima à Usina Hidrelétrica de Três Marias quanto à composição de espécies, abundância e suas relações com variáveis abióticas. Foram realizadas 18 coletas com redes de emalhar no canal de fuga da usina, durante um ano hidrológico, em seis horários ao longo de 24 h, quando também foram obtidos dados abióticos. Foi observado que no canal de fuga da usina, Characiformes apresentou uma maior riqueza, enquanto os Siluriformes foram os mais abundantes. Das 33 espécies coletadas seis eram migradoras. A comunidade de peixes foi significativamente diferente entre os meses de seca e chuva, com maior abundância no período de chuva. Neste período, a frequência das espécies migradoras na comunidade subiu de 0,7 para 18,3%, sendo que Pimelodus maculatus representou 14,5% da abundância. A maior captura de Characiformes se deu no período da manhã com pico de atividade de 5:00 às 9:00 h. Os Siluriformes apresentaram maior atividade à noite, principalmente das 17:00 às 21:00 h. Conciliar manobras que oferecem mais riscos para os peixes com o período de menor abundância no canal de fuga, ou seja, nos meses de baixa pluviosidade, pode ser considerada medida para reduzir o impacto da operação sobre a comunidade à jusante.
Introduction
Hydroelectric generation accounts for more than 76% of the energy produced in Brazil (EPE, 2010). Damming rivers to implement hydroelectric dams is considered as a discontinuity in the concept of the river continuum (Vannote et al., 1980; Stanford & Ward, 2001), causing a series of changes in the environment that can affect aquatic fauna, especially fish (Barletta et al., 2010). Downstream from the dam, there are changes affecting fluctuations in natural discharge and control of the flood regime (Baxter, 1977; Freeman et al., 2001; Gehrke & Harris, 2001; Zdankus et al., 2008). Damming interrupts fish migration (Dugan et al., 2010), causes gas supersaturation due to the incorporation of atmospheric gases by spillways and turbines (Schisler & Bergersen, 1999; Urban et al., 2008), affecting certain fish species due to maneuvers of turbines and spillways (Andrade et al., 2012; Schilt, 2007) and can increase levels of predation as a result of the high density of fish that accumulate immediately downstream (Agostinho et al., 2007). Dams also affect downstream populations by altering water quality and habitat conditions (Balciunas & Zdankus, 2007; García et al., 2011), with consequences for both resident and migratory fishes (Lessard & Hayes, 2003).
It is believed that hydrodynamic conditions in the stretches immediately downstream from a dam may attract fish onto the tailrace (Schilt, 2007; Agostinho et al., 2007). However, other environmental factors that also may determine species distributions remain poorly studied. In most hydroelectric dams, it is common to see large concentrations of fish aggregation downstream from the dam, in the tailrace (Agostinho et al., 2007). The presence of these aggregation is a concern for the electric energy segment because many power dams operations put ichthyofauna at risk (Ferguson et al., 2006; Schilt, 2007; Loures, 2009).
Several studies that address changes in fish community downstream from power dams, assess: differences from before and after damming (Mérona et al., 2005; Penczak & Kruk, 2005; Mérona et al., 2010), the processes of habitat fragmentation that interrupt migratory routes (Oldani & Baigún, 2002; Pelicice & Agostinho, 2008; Godinho & Kynard, 2009) and the effects of discharge and flood pulse on the communities (Agostinho et al., 2004; Bednarek & Hart, 2005; Welcomme et al., 2006; Godinho et al., 2007; Bailly et al., 2008). However, few studies have investigated the fish community in the tailraces of dams and how their knowledge can help reduce the impacts generated by its operation (Ferguson et al., 2006; Schilt, 2007; Nestler & Goodwin, 2001). Thus, we evaluate temporal variations in fish community immediately downstream from the Três Marias Hydroelectric Dam on the species composition and abundance and their relationships with abiotic variables.
Material and Methods
Study area
The São Francisco River basin is the third largest in Brazil, covering an area of 638,576 km2, which corresponds to 7.5% of the country and 40% of the Minas Gerais State. The basin is fully contained within Brazilian territory and is of great importance in the country, not only due to the volume of water transported across a semiarid region, but also because of the hydroelectric potential and its historical and economic contribution to regional development (ANA, 2009). The São Francisco River is 2,863 km long, originating in the Serra da Canastra in the State of Minas Gerais, flows in a south-to-north direction across the States of Bahia and Pernambuco, where the course turns to the southeast. The river's mouth to the Atlantic Ocean is located between Alagoas and Sergipe. The estimated hydroelectric potential of the hydrogeographic region is approximately 25,320 MW, and 10,380 MW of this potential is already installed, supplying energy for 16% of the country (MMA, 2006).
Currently, there are 18 dams in operation in the São Francisco River basin, and nine of them are located in the São Francisco River itself. The Três Marias Hydroelectric Dam (TMD), officially called "Bernardo Mascarenhas", is located on the São Francisco River (18°12'49"S 45°15'42"W) in the municipality of Três Marias (Fig. 1). The dam is 75 m high and 2,700 m long. It was inaugurated in 1961, and started the operation in July 1962. The installed capacity is 396 MW, with six generation units of 65 MW each. The nominal drop is 46.1 m, and energy is generated by Kaplan turbines with maximum discharge of 150m3/s. The spillway is of the controlled-surface type, with seven gates and a maximum capacity of 8,700 m3/s (Cemig, 2006). The turbinate water is taken from the hypolimnion of the reservoir (50 - 60 m deep), which has a lower temperature and dissolved oxygen level than the surface water. The temperature of the river water downstream from the dam can vary from 22.6 to 28.5°C, and the dissolved oxygen content can range from 1.89 mg/l in the summer to 7.76 mg/l in the winter. Turbidity is lower during periods of drought and higher in periods of heavy precipitation, ranging from 0 to 1000 NTU (Sampaio & López, 2003). The patterns of precipitation and discharge downstream from the dam over the past ten years (Mar 2001 to Mar 2011) show a regularity in the discharge over time and a concentration of precipitation from November to March (Fig. 2).
Fish collection
We sampled fishes monthly from April 2010 to September 2010 and fortnightly from October 2010 to March 2011, with a total of 18 samples. At each sample we set gillnets in the backwater of the tailrace over 24 hours and checked for fishes six times at 1:00, 5:00, 9:00, 13:00, 17:00, and 21:00 h. We used 10 m gill nets with mesh sizes varying from 3 to 16 cm (opposite knots length), with an approximately effort of 160 m². Fishes were identified and a group of voucher individuals was deposited at the Ichthyological Collection of the Federal University of Lavras (CIUFLA).
Abiotic data sampling
Limnological and hydrological data were used to assess the relationships between fish abundance and richness and abiotic factors in the TMD tailrace. Therefore, the acquisition of data followed the days and times of fish sampling in the backwater of the tailrace. We used a multiparameter probe (Horiba U10) to acquire water parameters, pH, dissolved oxygen (DO) and temperature (°C). Data regarding precipitation and discharge (turbinate and spill) for the study period were obtained from Cemig Geração e Transmissão S.A.
Data analysis
We evaluated fish community by an ordination based on species abundance, running a nMDS (nonmetric multidimensional scaling) test in the statistical program Past, using the Bray-Curtis index. The difference between dry and rainy seasons was tested by ANOSIM (analysis of similarity), and the contribution of each species to the dissimilarity was assessed by a SIMPER (Similarity Percentage) test.
In order to investigate the relationships between abiotic variables and the richness and abundance of fish, throughout the study period, we performed multiple regressions. The abundance of the ten most common species was also tested separately. It was necessary to normalize the abundance of each species using log10 (n+1), where n was the number of individuals of a species. We proceeded the statistical analysis (α=0.05) using Statistica 10.0 (StatSoft, 2010).
Results
We sampled 995 individuals belonging to 4 orders, 11 families, 24 genera and 33 species (Table 1). The orders Characiformes (29.8%) and Siluriformes (69.7%) together represented 99.5% of the total capture. Our sample included six migratory species and one exotic to the São Francisco River basin, Metynnis maculatus.
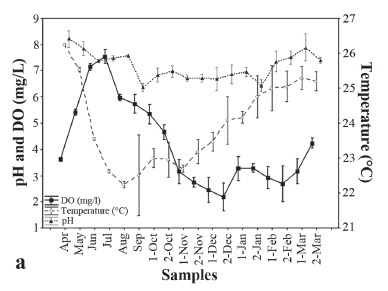
In the São Francisco River, downstream from the dam, the mean DO ranged from 2.2 to 7.6 mg/l over the course of a hydrological year, with higher values during the winter (Fig. 3a). The pH varied between 6.4 and 8.2, with higher values during the fall. The temperature, as expected, showed the opposite trend, with lower values observed in the winter and a gradual increase toward its maximum in the summer. The temperature ranged from 22.3 to 26.3 oC (Fig. 3a).
During the study period, the total discharge of the TMD ranged from 420.0 to 2,677.0 m3/s, with a daily average of 544.0 m3/s (Fig. 3b). In the second half of March 2011, there was an average spill of 814.1 m3/s, which elevated the total discharge during that month. It was also necessary to open the spillway in other months, but with lesser discharges: May - 66 m3/s and December - 59 m3/s. The average turbinate water discharge in the twelve months of the study was 507.6 m3/s.
From April to October (dry season), the average monthly accumulated precipitation in the region of the TMD was 30.6 mm, and during July and August, there was no rain in the area. During the period considered to be the wet season (November to March), the average monthly accumulated precipitation was 212.0 mm, with the highest precipitation in late December and early January (Fig. 3b).
The analysis of nMDS and ANOSIM (p = 0.0004) indicated that fish community was significantly different between dry (April to September) and wet (November to March) seasons (Fig. 4). The species that contributed most to the dissimilarity observed were Franciscodoras marmoratus, Pimelodus maculatus and Leporinus taeniatus, all with higher abundance during the wet season (Table 2).
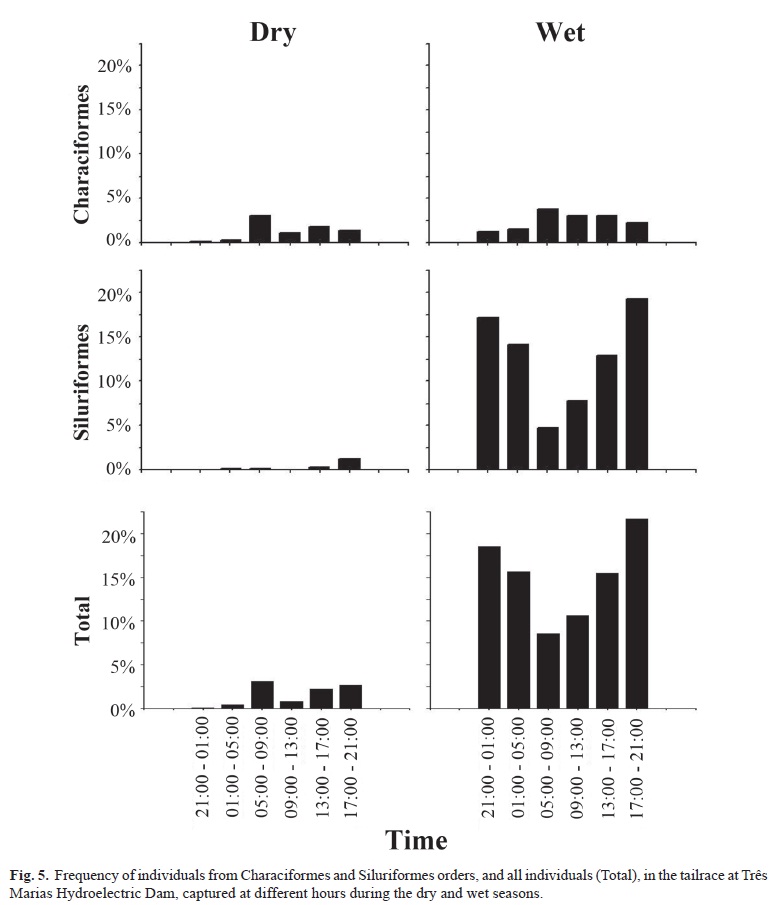
Considering the groups formed by nMDS analysis and the periods of fishes activity, we could observe that, during the dry season fish were more captured during the day, between 5:00 and 9:00 h (Fig. 5). Characiformes fishes were the most representative in this time while Siluriformes were captured in lower abundances and were restricted to the interval between 13:00 and 21:00 h. During the wet season, the abundance of fish was higher at all times when compared to the dry season. The Characiformes remained with a peak in captures during the day from 5:00 to 9:00 h, while the Siluriformes, most abundant in this season, were more captured at night, especially from 17:00 to 21:00 h (Fig. 5).
We did not find any correlation between the measured water physical and chemical parameters and the total abundance of fish, but total discharge (turbinate and spill) and precipitation explained 73% of the variance observed in the number of individuals captured (Table 3). Species richness was explained by total discharge and precipitation (Table 3). Of the 10 most abundant species, only for Hypostomus cf. margaritifer, Leporinus taeniatus, Pimelodus pohli and Pseudopimelodus charus we did not find significant relationships between abundance and abiotic variables tested (Table 3). The abundance of A. fasciatus was negatively related with the variation of DO (r2=0.22). Temperature explained the variance in abundance of Franciscodoras marmoratus (44%) and of Serrasalmus brandtii (37%). Both precipitation and temperature were positively correlated with Pimelodus maculatus abundance, and pH was negatively correlated with it. Discharge explained 80% of the variance in Prochilodus costatus abundance and, together with precipitation, it also explained 63% of the abundance of Rhinelepis aspera.
Discussion
Since the late XVIII century, ichthyofauna surveys of São Francisco River basin has been carried out (Sato & Godinho, 1999). However, for the region of Três Marias, the most important work on fish systematic was not published until 1984, where 73 species were recorded for the region of Três Marias and 132 for the entire São Francisco River Basin (Britski et al., 1984), becoming a landmark for studies of fish in the basin and considered indispensable to all those interested in studying the fish fauna of the region (Alves et al., 2011). Currently in the São Francisco River basin, a total of 205 species of native fish have been recorded, and the orders Characiformes and Siluriformes account for 74% of species richness (Alves et al., 2011). In the TMD tailrace, although Characiformes had the highest species richness, Siluriformes were the most abundant. This composition was also observed close to the dam, downstream Samuel Hydroelectric Dam on the Jamari River in the State of Rondônia. After the formation of the reservoir, there was a reduction in the abundance of Characiformes, and Siluriformes became prevalent, mainly due to an increase in the abundance of a Pimelodus species (Santos, 1995). During the period of highest abundance in the tailrace, we observed fishermen capturing "curimbas" (Prochilodus spp.) with cast nets, whereas the gill nets we applied only captured 18 individuals. Thus, the selectivity of the fishing gear used may be underestimated the fish community, once the capture depends on the activity of the fishes as well as body size (Hubert, 1996).
In Três Marias, the São Francisco and Parnaíba Valley Development Company (Companhia de Desenvolvimento do Vale São Francisco e Parnaíba CODEVASF) conducts regular monitoring with gill nets in two stretches of the river downstream from the TMD. The nearest sample point is more than 1 km away from the tailrace. In 2010, these monitoring results showed a fish community composition different from that observed in the tailrace in our study, with a predominance of Characiformes in both richness and abundance (CODEVASF, unpublished data). This finding could indicate that, for studies investigating the direct impacts of a power dam on fish community (such as injuries caused by turbines and entrapment in the draft tube), samples far from the tailrace may not represent the community most susceptible to those impacts.
We observed that lower values of dissolved oxygen in tailrace water occurred during warmer months, most likely due to thermal stratification of the reservoir and a bottom water uptake to electricity generation by the dam (Sampaio & López, 2003). In addition, the increased water temperature during this period may have decreased the solubility of oxygen. Low DO concentrations can affect the swimming performance of fish and reduce fertility rates, but the limit is species-specific (Portz et al., 2006). The water temperature in the tailrace begins to rise in November, with highs in the summer. It is known that the water temperature increase can be a trigger for some species to initiate upstream migration toward their spawning sites (Lowe-McConnell, 1999). In general, most natural freshwater systems have a pH ranging from 5.0 to 10.0 (Portz et al., 2006), similar to the values found in TMD tailrace.
Fish migrate to find better environmental conditions, shelter, food and appropriate locations to finish their development and to reproduce (Lowe-McConnell, 1999). In dams that do not have fish passages, such as TMD, fishes can concentrate at the base of the dam trying to reach upstream stretches, during juvenile migration (known as "arribação") or spawning migration (known as "piracema") (Agostinho et al., 2007; Godinho & Kynard, 2006). We did not observe an increase in abundance from April to July, characteristic of "arribação" (Godinho & Kynard, 2009). However, the highest proportion of Characiformes captured in these months, it was also found near the dam in 2007, when the most pronounced juvenile migration in the last four years occurred in the São Francisco River (Rodrigues, 2009). During the wet season (November to March), which coincides with the breeding season of many species of the São Francisco River (Alves et al., 2011), the highest abundance of fish was observed in the tailrace. During this period, the frequency of migratory species in the community rose from 0.7 to 18.3%, and P. maculatus accounted for 14.5% of this abundance.
The difference in the composition of fish community between dry and wet months was a result of an increase in the abundance of two species of Siluriformes: Franciscodoras marmoratus and Pimelodus maculatus. The latter species is the one most impacted by turbine maneuvers and draft tube dewatering at TMD (Andrade et al., 2012). In both cases there is a high risk of fish mortality or injuries. Pimelodus maculatus is a species with high swimming performance when compared to temperate fish, and shows preference for swimming near the bottom of the channel and walls of the dam structure. This behavior probably contributes to these mortality events, once fish can access the draft tube more easily (Santos et al., 2008). Thereby, as a measure to reduce the impact on the community, we recommend conciliate activities more risky to the fish with periods of lowest abundance in the tailrace (i.e., in the months of low precipitation).
Siluriformes are a nocturnal group (Burgess, 1989), whereas Characiformes present a more diverse pattern of activities with a diurnal peak (Pompeu & Martinez, 2006). These patterns were confirmed by the present study. During the dry season, the Characiformes were more abundant than Siluriformes, and the higher captures of fish occurred in the morning, with peak activity from 5:00 to 9:00 h. In the wet season, the abundance of fish increased at all times, with a predominance of Siluriformes, especially at night from 17:00 to 21:00 h. The TMD does not program neither execute maneuvers that may represent risk to the fish, such as turbine dewatering and stop/startup, at night (Cemig, unpublished data). Considering our results, this measure could reduce the risk of fish mortality, especially during the wet season. During this season, if a turbine stops at night, when the abundance and activity of fishes in the tailrace is higher, the probability of fish enter into the draft tube increase. Thus, the risk of mortality would be higher leading to greater impacts on fish community. We suggest that complementary studies should be developed in order to evaluate the presence of fishes not only in the tailrace, but also inside the draft tube during day and night.
The total discharge and precipitation explained more than 70% of the variance observed in the number of individuals sampled in the tailrace. Furthermore, these variables explained the change in richness over time. Fish exhibit a spectrum of life cycle stages associated with the seasonality of tropical waters. In tropical rivers, seasonal differences are mainly driven by changes in the water level (Lowe-McConnell, 1999). Additionally, precipitation and temperature may trigger spawning migrations (Parkinson et al., 1999). Despite the control of river discharge due to regulation by the TMD, it was observed that 80% of the abundance of the migratory species Prochilodus costatus was explained by variation in the total discharge of the dam. Along with precipitation, the discharge also explained 63% of the variance in the abundance of Rhinelepis aspera. Although this species is considered migratory in the Paraná river basin (Agostinho et al., 1986; Agostinho et al., 1995), the characteristics of egg and larval individuals from the São Francisco River indicate that R. aspera does not require long reproductive migrations, like Pimelodus maculatus (Sato et al., 2003, Sato & Godinho, 2003). The increase in abundance in response to precipitation and discharge observed in this study, could be a sign that this species also migrates along the São Francisco River.
Three species responded to temperature change: Franciscodoras marmoratus, Serrasalmus brandtii and Pimelodus maculatus. Abundances of P. maculatus were also positively correlated with precipitation and negatively correlated with pH. The importance of precipitation to increase the abundance of Siluriformes downstream from dams has been previously documented (Pompeu & Martinez, 2006), as well as the role of temperature as a trigger for breeding (Parkinson et al., 1999; Sato et al., 2005). However, the role of pH is not supported in the literature. As P. maculatus and R. aspera, the eggs and larvae of F. marmoratus indicate that this species does not require long reproductive migrations (Sato et al., 2003). More studies are needed to assess whether the increase in water temperature is indeed a trigger for this species to initiate reproductive migrations. The drop in DO levels during the wet season explained the abundance of Astyanax fasciatus in the tailrace. In general, fish tend to avoid sites with a low DO level (Portz et al., 2005), so it would be interesting evaluate if the abundance increase was related to environmental stimuli or some other biological factor that we did not assess.
The system of production and transmission of electric energy in Brazil can be considered unique in the world due to its size and distinct characteristics. The National Interconnected Power System (Sistema Interligado Nacional SIN) is formed by companies from five administrative regions: South, Southeast, Midwest, Northeast and part of the North. Only 3.4% of the electricity capacity of production in Brazil occurs outside of the SIN, in small isolated systems located mainly in the Amazon region. The Electric System National Operator (Operador Nacional do Sistema Elétrico ONS) is responsible for coordinating and controlling the operation of existing electric facilities in the SIN, with surveillance and regulation by the Brazilian Electricity Regulatory Agency (Agência Nacional de Energia Elétrica ANEEL) (ONS, 2011). In practice, this structural organization means that, the power companies must submit operational maneuvers programming to ONS approval. Thus restrictions to dam operation concerning different aspects (electromechanical, electric, civil, social and multiple-use, discharge control and environment) should be analyzed jointly by the electric sector and regulatory agencies (Cavallari, 2009). Operate a power dam with less risk of mortality to downstream fish community is a challenge faced by the hydroelectric companies. In order to determine better ways to operate with less risks to fishes, besides the knowledge of fish community near the dam, it is necessary to consider a number of other restrictions and legal procedures to get the approval from the ONS for a change in generation.
Nevertheless, information regarding community and behavior of fish downstream dams may improve the arguments between environmental and operational areas from a power company. Moreover together with regulatory agencies from the electric sector, such information can help define procedures with better cost-benefit operational-environmental relations.
Acknowledgements
We thank the fisherman Waldir Alves dos Santos and the biologists Ivo Prado, Igor Boratto and Francisco de Andrade, for their support in data collection. We also thank the environmental analyst Renato Junio Constâncio and the Três Marias Hydroelectric Dam operation and maintenance team for their assistance in field work, and the Peixe Vivo Program of CEMIG for providing financial support.
Literature Cited
Submitted March 14, 2012
Accepted August 16, 2012
- Agostinho, A. A., G. Barbieri, J. R. Verani & C. S. Agostinho. 1986. Ciclo reprodutivo e primeira maturação de Rhinelepis aspera (Agassiz 1829), (Teleostei -Loricariidae) no rio Paranapanema. Revista Unimar, 8: 17-27.
- Agostinho, A. A., L. C. Gomes & F. M. Pelicice. 2007. Ecologia e Manejo de Recursos Pesqueiros em Reservatórios do Brasil. Eduem, 501p.
- Agostinho, A. A., L. C. Gomes, S. Veríssimo & E. K. Okada. 2004. Flood regime, dam regulation and fish in the Upper Paraná River: effects on assemblage attributes, reproduction and recruitment. Reviews in Fish Biology and Fisheries, 14: 11-19.
- Agostinho, A. A., Y. Matsumura, E. K. Okada & K. Nakatani. 1995. The catfish, Rhinelepis aspera (Teleostei; Loricariidae), in the Guaíra region of the Paraná river: an example of population estimation from catch-effort and tagging data when emigration and immigration are high. Fisheries Research, 23: 333-344.
- Alves C. B. M., F. Vieira & P. S. Pompeu. 2011. Ictiofauna da bacia hidrográfica do rio São Francisco. In: Ministério do Meio Ambiente. (Org.). Diagnóstico do macrozoneamento ecológico-econômico da bacia hidrográfica do rio São Francisco. Ministério do Meio Ambiente, Brasília, 226-241.
- ANA -Agencia Nacional de Água. 2009. Água: edição comemorativa do ano 10 da Agencia Nacional de Água. Brasília, 264p.
- Andrade, P. M. & F. M. S. Braga. 2005. Reproductive seasonality of fishes from a lotic stretch of the Grande river, High Paraná river basin, Brazil. Brazilian Journal of Biology., 65: 387-394.
- Andrade, F., I. G. Prado, R. C. Loures & A. L. Godinho. 2012. Evaluation of techniques to protect tailrace fishes during turbine maneuvers at Três Marias Dam, Brazil. Neotropical Ichthyology, 10(4): 723-730.
- Bailly, D., A. A. Agostinho & H. I. Suzuki. 2008. Influence of the flood regime on the reproduction of fish species with different reproductive strategies in the Cuiabá River, Upper Pantanal, Brazil. River Research and Applications, 24: 1218-1229.
- Balciunas, P. & N. Zdankus. 2007. Harmonization of hydropower plant with the environment. Renewable and Sustainable Energy Reviews, 11: 1260-1274.
- Barletta, M., A. J. Jaureguizar, C. Baigun, N. F. Fontoura, A. A. Agostinho, V. M. F. Almeida-Val, A. L. Val, R. A. Torres, L. F. Jimenes-Segura, T. Giarrizzo, N. N. Fabre, V. S. Batista, C. Lasso, D. C. Taphorn, M. F. Costa, P. T. Chaves, J. P. Vieira & M. F. M. Correa. 2010. Fish and aquatic habitat conservation in South America: a continental overview with emphasis on neotropical systems. Journal of Fish Biology, 76: 2118-2176.
- Baxter, R.M. 1977. Environmental effects of dams and impoundments. Annual Review of Ecology and Systematics, 8: 255-283.
- Bednarek, A. T. & D. D. Hart. 2005. Modifying dam operations to restore rivers: ecological responses to tennessee river dam mitigation. Ecological Applications, 15: 997-1008.
- Britski, H. A., Y. Sato & A. B. S. Rosa. 1984. Manual de identificação de peixes da região de Três Marias: com chave de identificação para os peixes da bacia do rio São Francisco. 1Ş.ed. Codevasf, Brasília, 143p.
- Burguess, W. E. 1989. An atlas of freshwater and marine catfishes. Neptune City, TFH, 785p.
- Cavallari, A. 2009. Operação de reservatórios. Ação Ambiental, 12: 46-48.
- Cemig. 2006. Usinas da Cemig: 1952-2005, a história da eletricidade em Minas e no Brasil. Centro da Memória da Eletricidade no Brasil, Rio de Janeiro, 304p.
- Dugan, P. J., C. Barlow, A. A. Agostinho, E. Baran, G. F. Cada, D. Chen, I. G. Cowx, J. W. Ferguson, T. Jutagate, M. Mallen-Cooper, G. Marmulla, J. Nestler, M. Petrere, R. L. Welcomme & K. O. Winemiller. 2010. Fish migration, dams, and loss of ecosystem services in the Mekong Basin. AMBIO, 39:344-348.
- EPE – Empresa de Pesquisa Energética. 2010. Balanço Energético Nacional 2010: Ano base 2009. Rio de Janeiro, 276p.
- Ferguson, J. W., R. F. Absolon, T. J. Carlson & B. P. Sandford. 2006. Evidence of delayed mortality on juvenile Pacific Salmon passing through turbines at Columbia River Dams. Transactions of the American Fisheries Society, 135: 139-150.
- Freeman, M. C., Z. H. Bowen, K. D. Bovee & E. R. Irwin. 2001. Flow and habitat effects on juvenile fish abundance in natural and altered flow regimes. Ecological Applications, 11: 179-190.
- García, A., K. Jorde, E. Habit, D. Caamaño & O. Parra. 2011. Downstream environmental effects of dam operations: changes in habitat quality for native fish species. River Research and Applications, 27: 312-327.
- Gehrke, P. C. & J. H. Harris. 2001. Regional-scale effects of flow regulation on lowland riverine fish communities in New South Wales, Australia. Regulated Rivers: Research & Management, 17: 369-391.
- Godinho, A. L. & B. Kynard. 2006. Migration and spawning of radio-tagged zulega Prochilodus argenteus in a dammed Brazilian river. Transactions of the American Fisheries Society, 135: 811-824.
- Godinho, A. L., B. Kynard & C. B. Martinez. 2007. Supplemental water releases for 18 fisheries restoration in a Brazilian floodplain river: a conceptual model. Rivers Research and Applications, 3: 1-16.
- Godinho, A. L. & B. Kynard. 2009. Migratory fishes of Brazil: life history and fish passage needs. River Research and Applications, 25: 702-712.
- Hubert, W. A. 1996. Passive capture techniques. In: Murphy, B.R. & Willis D. W. (Eds.) Fisheries Techniques 2nd edition. American Fisheries Society, Bethesda, Maryland, 752p.
- Lessard, J. L. & D. B. Hayes. 2003. Effects of elevated water temperature on fish and macroinvertebrate communities below small dams. River Research and Applications, 19: 721-732.
- Loures, R. C. F. 2009. A compatibilização da operação de usinas com a ictiofauna. Ação Ambiental, 12: 24 -28.
- Lowe-McConnell, R. H. 1999. Ecologia de Comunidades de Peixes Tropicais. EDUSP, São Paulo, SP. 535p.
- Mérona, B., A. A. Juras, G. M. Santos & I. H. A. Cintra. 2010. Os peixes e a pesca no baixo Rio Tocantins: vinte anos depois da UHE Tucuruí, 208p.
- Mérona, B., R. Vigouroux & F. L. Tejerina. 2005. Alteration of fish diversity downstream from Petit-Saut Dam in French Guiana: Implication of ecological strategies of fish species. Hydrobiologia, 551:33-47.
- MMA - Ministério do Meio Ambiente. 2006. Caderno setorial de recursos hídricos: geração de energia hidrelétrica. Secretaria de Recursos Hídricos, Brasília, 112p.
- Nestler, J. M. & A. Goodwin. 2001. Simulating fish movement in hydropower reservoirs and tailwaters. Hydro Review ,1-4.
- Oldani, N. O. & C. R. M. Baigún. 2002. Performance of a fishway system in a major South American dam on the Parana river (Argentina - Paraguai). River Research and Applications, 18: 171-183.
- ONS - Operador Nacional do Sistema Elétrico. 2011. Available at: http://www.ons.org.br/conheca_sistema/o_que_e_sin.aspx Accessed May 1, 2011.
» link - Parkinson, D., J. Philippart & E. Baras. 1999. A preliminary investigation of spawning migrations of grayling in a small stream as determined by radio-tracking. Journal of Fish Biology, 55: 172-182.
- Penczak, T. & A. Kruk. 2005. Patternizing of impoundment impact (1985-2002) on fish assemblages in a lowland river using the Kohonen algorithm. Journal of Applied Ichthyology, 21, 169-177.
- Pelicice, F. M. & A. A. Agostinho. 2008. Fish-passage facilities as ecological traps in large Neotropical rivers. Conservation Biology, 22: 180-188.
- Pompeu, P. S. & C. B. Martinez. 2006. Variações temporais na passagem de peixes pelo elevador da Usina Hidrelétrica de Santa Clara, rio Mucuri, leste brasileiro. Revista Brasileira de Zoologia, 23: 340-349.
- Portz, D. E., C. M. Woodley & J. J. Cech Jr. 2006. Stress-associated impacts of short-term holding on fishes. Reviews in Fish Biology and Fisheries, 16: 125-170.
- Rodrigues, R. R. 2009. Sucesso reprodutivo de peixes migradores em rios barrados em Minas Gerais: influência da bacia de drenagem e das cheias. Unpublished Master Dissertation, Universidade Federal de Minas Gerais, Belo Horizonte, 56 p.
- Sampaio, E. V. & C. M. López. 2003. Limnologias física, química e biológica da represa de Três Marias e do São Francisco. Pp 71-92.In: Godinho H. P, Godinho A. L. (Eds.). Águas, peixes e pescadores do São Francisco das Minas Gerais. PUC Minas, Belo Horizonte.
- Santos, G. M. 1995. Impacto da Hidrelétrica Samuel sobre as comunidades de peixes do rio Jamari (Rondônia, Brasil). Acta Amazonica, 25: 247-280
- Santos, H. A., P. S. Pompeu, G. S. Vicentini & C. B. Martinez. 2008. Swimming performance of the freshwater neotropical fish: Pimelodus maculatus Lacepède, 1803. Brazilian Journal of Biology, 68: 433-439.
- Sato, Y., N. Bazzoli, E. Rizzo, M. B. Boschi & M. O. T. Miranda. 2005. Influence of the Abaeté river on the reproductive success of the neotropical migratory teleost Prochilodus argenteus in the São Francisco River, downstream from the Três Marias Dam, Southeastern Brazil. River Research and applications 21: 939-950.
- Sato, Y., N. Fenerich-Verani, A. P. O. Nuñer, H. P. Godinho & J. R. Verani. 2003. Padrões reprodutivos de peixes da bacia do São Francisco. In: Godinho HP, Godinho AL (eds) Águas, peixes e pescadores do São Francisco das Minas Gerais. PUC Minas, Belo Horizonte, 468p.
- Sato, Y. & H. P. Godinho. 1999. Peixes da Bacia do Rio São Francisco. In: Lowe-McConnell, R. H. Ecologia de Comunidades de Peixes Tropicais. EDUSP, São Paulo, SP. 535p
- Sato, Y. & H. P. Godinho. 2003. Migratory fishes of the São Francisco River. Pp. 195-232. In: Carolsfeld J., Harvey B., Ross C., Baer A. (Eds.) Migratory fishes of South America. World Fisheries Trust/The World Bank/International Development Research Centre, Ottawa.
- Schilt, C. R. 2007. Developing fish passage and protection at hydropower dams. Applied Animal Behaviou Science, 104: 295-325.
- Schisler, G. J. & E. P. Bergersen., 1999. Identification of gas supersaturation sources in the Upper Colorado River, USA. Regulated River: Research & Management, 15: 301-310.
- Stanford, J. A. & J. V. Ward. 2001. Revisiting the serial discontinuity concept. Regulated Rivers: Research & Management, 17: 303-310.
- Urban, A. L., J. S. Gulliver & D. W. Johnson. 2008. Modeling total dissolved gas concentration downstream of spillways. Journal of Hydraulic Engineering, 134: 550.
- Vannote, R. L., G. W. Minshall, K. W. Cummins, J. R. Sedell & C. E. Cushing. 1980. The river continuum concept. Canadian Journal of Fisheries and Aquatic Sciences, 37:130-137.
- Welcomme, R. L., K. O. Winemiller & I. G. Cowx. 2006. Fish environmental guilds as a tool for assessment of ecological condition of rivers. River Research and Applications, 22: 377-396.
- Zdankus, N, S. Vaikasas & G. Sabas. 2008. Impact of a hydropower plant on the downstream reach of a river. Journal of Environmental Engineering and Landscape Management, 16: 128-134.
Publication Dates
-
Publication in this collection
31 Oct 2012 -
Date of issue
Oct 2012
History
-
Received
14 Mar 2012 -
Accepted
16 Aug 2012