
Abstracts
Developing survey strategies for threatened species is often limited by access to sufficient individuals to acquire information needed to design appropriate conservation measures. The goliath grouper (Epinephelus itajara) is a large reef fish, globally classified as critically endangered. In Brazil, fishing has been prohibited since 2002. Herein we investigated habitat use and abundance of E. itajara in Brazil drawn from a participative survey, which engaged volunteer divers in data-collection. A total of 188 reports were recorded between 2005 to 2011 throughout 13 Brazilian states. Our results revealed that habitat type influenced the relationship between total length and depth of occurrence of E. itajara. We observed a significant difference between habitats, artificial presented a nonlinear and natural an asymptotic relationship. This study underscores the importance of developing conservation measures addressing specific habitats and locations to enhance population recovery. Artificial structures (shipwrecks and manmade reefs) are critical habitats that must be considered as highly sensitive areas to E. itajara strategic conservation policies. We suggest the creation and implementation of marine protected areas, as key strategies for E. itajara conservation, especially for artificial habitats and aggregation sites.
O desenvolvimento de estratégias de pesquisa para espécies ameaçadas é frequentemente limitado pelo acesso a indivíduos suficientes para adquirir informações necessárias para elaborar medidas de conservação adequadas. Dentre essas espécies, o mero (Epinephelus itajara) é um peixe recifal de grande tamanho, classificado globalmente como criticamente ameaçado. No Brasil, a pesca é proibida desde 2002. Investigou-se o uso de hábitat e abundância de E. itajara no Brasil através de uma pesquisa participativa, que engajou mergulhadores voluntários na coleta dos dados. Um total de 188 relatos foram registrados entre 2005 a 2011 em 13 estados brasileiros. Os resultados revelaram que o tipo de hábitat influenciou a relação entre o comprimento total e profundidade de ocorrência de E. itajara. Observou-se uma diferença significativa entre hábitats: hábitat artificial apresentou uma relação não linear e hábitat natural uma relação assintótica. Este estudo ressalta a importância do desenvolvimento de medidas de conservação abordando hábitats e locais específicos visando a recuperação populacional. Estruturas artificiais (naufrágios e recifes feitos pelo homem) são hábitats críticos que devem ser considerados como áreas relevantes para as estratégias políticas conservação de E. itajara. Sugere-se a criação e implementação de áreas marinhas protegidas como uma estratégia fundamental para a conservação de E. itajara, especialmente em hábitats artificiais e locais de agregação.
Introduction
Surveying rare and threatened species is often limited by access to sufficient individual records, challenging the build-up of comprehensive data sets needed to design appropriate conservation measures (Thompson, 2004Thompson, W. L . 2004. Sampling rare or elusive species. Island Press: Washington, D.C.; Wiley & Simpfendorfer, 2010Wiley, T. & C. Simpfendorfer. 2010. Using public encounter data to direct recovery efforts for the endangered smalltooth sawfish Pristis pectinata. Endangered Species Research, 12: 179-191.). This is particularly the case for species that live in marine habitats where they can be difficult to locate and capture (Carlson et al., 2013Carlson, J. K., S. J. B. Gulak, C. A. Simpfendorfer, R. D. Grubbs & J. G. Romine. 2013. Movement patterns and habitat use of smalltooth sawfish, Pristis pectinata, determined using pop-up satellite archival tags. Aquatic Conservation: Marine and Freshwater Ecosystems, DOI: 10.1002/aqc.2382.
https://doi.org/10.1002/aqc.2382...
). One approach to overcoming these challenges is the engagement of volunteers to collect data (Foster-Smith & Evans, 2003Foster - Smith, J. & S. M. Evans. 2003. The value of marine ecological data collected by volunteers. Biological Conservation, 113: 199-213.; Holt et al., 2013Holt, B. G., R. Rioja-Nieto, M. A. MacNeil, J. Lupton & C. Rahbek. 2013. Comparing diversity data collected using a protocol designed for volunteers with results from a professional alternative. Methods in Ecology and Evolution, 4: 383-392.).
Participative ichthyologic research using volunteer diver databases has been widely applied over the past two decades (e.g., Darwal & Dulvy, 1996Darwal, W. R. T. & N. T. Dulvy. 1996. An evaluation of the suitability of non-specialist volunteer researchers for coral reef fish surveys. Mafia Island, Tanzania - A case study. Biological Conservation, 78: 223-231.; Auster et al., 2005Auster, P. J., B. X. Semmens & K. Barber. 2005. Pattern in the co-occurrence of fishes inhabiting the coral reefs of Bonaire, Netherlands Antilles. Environmental Biology of Fishes, 74: 187-194.; Stallings, 2009Stallings, C. 2009. Fishery-independent data reveal negative effect of human population density on caribbean predatory fish communities. PLoS ONE, 4: e5333.; Ruttenberg et al., 2012Ruttenberg, B. I., P. J. Schofield, J. L. Akins, A. Acosta, M. W. Feeley, J. Blondeau, S. G. Smith & J. S. Ault. 2012. Rapid invasion of Indo-Pacific lionfishes (Pterois volitans and Pterois miles) in the Florida Keys, USA: evidence from multiple pre-and post-invasion data sets. Bulletin of Marine Science, 88: 1051-1059.). Recreational divers represent a potentially large community of skilled and often highly motivated volunteers (Hussey et al., 2013Hussey, N. E., N. Stroh, R. Klaus, T. Chekchak & S. T. Kessel. 2013. SCUBA diver observations and placard tags to monitor grey reef sharks, Carcharhinus amblyrhynchos, at Sha'ab Rumi, The Sudan: assessment and future directions. Journal of the Marine Biological Association of the United Kingdom, 93, 299-308.). This approach has proven to be valuable in structuring consistent large-scale geographic databases encompassing marine system properties such as diversity and dynamics at multiple scales (Goffredo et al., 2010Goffredo, S., F. Pensa, P. Neri, A. Orlandi, M. S. Gagliardi, A. Velardi, C. Piccinetti & F. Zaccanti. 2010. Unite research with what citizens do for fun: ''recreational monitoring'' of marine biodiversity. Ecological Applications, 20: 2170-2187.; Holt et al., 2013Holt, B. G., R. Rioja-Nieto, M. A. MacNeil, J. Lupton & C. Rahbek. 2013. Comparing diversity data collected using a protocol designed for volunteers with results from a professional alternative. Methods in Ecology and Evolution, 4: 383-392.). However, the use of volunteers is often contested on the grounds that the information collected will be unreliable as a result of either insufficient training or lack of consistency derived from a large numbers of observers (Darwal & Dulvy, 1996Darwal, W. R. T. & N. T. Dulvy. 1996. An evaluation of the suitability of non-specialist volunteer researchers for coral reef fish surveys. Mafia Island, Tanzania - A case study. Biological Conservation, 78: 223-231.). By engaging with and accepting the limitations of recreational divers' observational data - i.e., size over estimation, replicate counting of individuals and divers disturbance on mobile animals (Hussey et al., 2013Hussey, N. E., N. Stroh, R. Klaus, T. Chekchak & S. T. Kessel. 2013. SCUBA diver observations and placard tags to monitor grey reef sharks, Carcharhinus amblyrhynchos, at Sha'ab Rumi, The Sudan: assessment and future directions. Journal of the Marine Biological Association of the United Kingdom, 93, 299-308.) - this approach has provided a number of publications about marine fishes, demonstrating the value of data collected by volunteer divers. Examples include a variety of studies on reef fish population abundance and distribution in the Caribbean and Italy (Goffredo et al., 2004Goffredo, A., C. Piccinetti & F. Zaccanti. 2004. Volunteers in marine conservation monitoring: A study of the distribution of seahorses carried out in collaboration with recreational scuba divers. Conservation Biology, 18: 1492-1503.; Ward-Paige et al., 2010Ward-Paige, C. A., C. Mora, H. K. Lotze, C. Pattengill-Semmens, L. McClenachan, E. Arias-Castro & R. A. Myers. 2010. Large-Scale Absence of Sharks on Reefs in the Greater- Caribbean: A Footprint of Human Pressures. PLoS ONE, 5: e11968.); seasonality of the manta ray Manta birostris in southeast Brazil (Luiz et al., 2009Luiz, O. J., A. P. Balboni, G. Kodja, M. Andrade & H. Marum. 2009. Seasonal occurrences of Manta birostris (Chondrichthyes: Mobulidae) in southeastern Brazil. Ichthyological Research, 56: 96-99.); evidences of recovery in goliath grouper Epinephelus itajara population in Florida U.S. (Koenig et al., 2011Koenig, C. C., F. C. Coleman & K. Kingon. 2011. Pattern of recovery of the goliath grouper Epinephelus itajara population in the southeastern US. Bulletin of Marine Science, 87: 891-911.; Frias-Torres, 2012Frias-Torres, S. 2012. Should the Critically Endangered Goliath grouper Epinephelus itajara be culled in Florida? Oryx, 1: 1-8.).
This paper undertakes a participative approach in the study of habitat use, depth of occurrence and abundance of E. itajara (Litchtenstein, 1822) (Perciformes: Epinephelidae) in Brazil. The species is the largest grouper in the Atlantic Ocean (>2 m total length - TL, >400 kg) (Bullock et al., 1992Bullock, L. H., M. D. Murphy, M. F. Godcharles & M. E. Mitchell. 1992. Age, growth and reproduction of jewfish Epinephelus itajara in the eastern Gulf of Mexico. Fisheries Bulletin, 90: 243-249.). They are sedentary and have a high degree of site fidelity (Eklund & Schull, 2001Eklund, A. M. & J. Schull. 2001. A stepwise approach to investigate the movement patterns and habitat utilization of goliath grouper, Epinephelus itajara, using conventional tagging, acoustic telemetry and satellite tracking. Pp. 189-216. In: Nielsen, J. L. (Ed.). Electronic tagging and tracking in marine fisheries. Amsterdam, Kluwer Academic Publishers.; Pina-Amargós & Gonzáles-Sansón, 2009Pina-Amargós, F. & G. González-Sansón. 2009. Movement of goliath grouper (Epinephelus itajara) in southeast Cuba: implications for its conservation. Endangered Species Research, 7: 243-247.). Adults inhabit both natural and artificial reefs, generally in depths less than 50 m (Bullock et al., 1992Bullock, L. H., M. D. Murphy, M. F. Godcharles & M. E. Mitchell. 1992. Age, growth and reproduction of jewfish Epinephelus itajara in the eastern Gulf of Mexico. Fisheries Bulletin, 90: 243-249.).
Historically, E. itajara was distributed throughout coastal and estuarine areas of the tropical and subtropical Atlantic (Heemstra & Randall, 1993Heemstra, P. C. & J. E. Randall. 1993. FAO species catalogue: Groupers of the world (Family Serranidae, subfamily Epinephelinae). An annotated and illustrated catalogue of the grouper, rockcod, hind, coral grouper and lyretail species known to date. FAO Fisheries Synopis, 16: 1-382.). However, historical records of drastic reductions in abundance have been observed throughout its range (Sadovy & Eklund, 1999Sadovy, Y. & A. Eklund. 1999. Synopsis of biological data on the Nassau Grouper, Ephinephelus striatus, and the Jewfish, E. itajara. NOAA Technical Report.; Gerhadinger et al., 2006Gerhardinger, L. C., R. Medeiros., R. C. Marenzi., A. A. Bertoncini & M. Hostim-Silva. 2006. Local Ecological Knowledge on the Goliath Grouper Epinephelus itajara (Teleostei: Serranidae) in Southern Brazil. Neotropical Ichthyology, 4: 441-450.; McClenachan, 2009McClenachan, L. 2009. Historical declines of goliath grouper populations in South Florida, USA. Endangered Species Research, 7: 175-181.). Although there is reduced survey effort in the East Atlantic, it is suggested to have become extinct in Africa by Craig et al. (2009)Craig, M. T., R. T. Graham, R. A. Torres, J. R. Hyde, M. O. Freitas, B. P. Ferreira, M. Hostim-Silva, L. C. Gerhardinger, A. A. Bertoncini & D. R. Robertson. 2009. How many species of goliath grouper are there? Cryptic genetic divergence in a threatened marine fish and the resurrection of a geopolitical species. Endangered Species Research, 7: 167-174.. Decreasing population abundances have resulted in the species being classified globally as critically endangered by the International Union for Conservation of Nature (IUCN, 2013IUCN. 2013. The IUCN Red List of Threatened Species. International Union for Conservation of Nature and Natural Resources. Available from: http://www.iucnredlist.org/details/195409/0 (10 Aug 2013).
http://www.iucnredlist.org/details/19540...
). In Brazil, it was the first marine fish subjected to a strict moratorium and has been protected from harvest since 2002 (Hostim-Silva et al., 2005Hostim-Silva M., A. A. Bertoncini, L. C. Gerhardinger & L. F. Machado. 2005. The 'Lord of the Rock's' conservation program in Brazil: the need for a new perception of marine fishes. Coral Reefs 24, 7.). One of the policy prescriptions attached to the moratorium has been the amalgamation of data to subsidize species' recovery and management solutions. With the purpose to provide conservation-relevant information, this paper reports on a Participative Survey Program (PSP) approach. Volunteer divers collected data on sightings of E. itajara, providing a description on its distribution along the Brazilian coast, considering natural and artificial habitats. Additionally, sites of aggregation events were mapped.
Material and Methods
Study area. The study was conducted through sporadic sightings of E. itajara along the Brazilian states from Maranhão to Santa Catarina (Fig. 1). The area presents a high richness of coastal environments (see Amaral & Jablonski, 2005Amaral, A. C. Z. & S. Jablonski. 2005. Conservation of Marine and Coastal Biodiversity in Brazil. Conservation Biology, 19: 625-631.). Sightings were carried out in tropical and subtropical coral and rocky reefs, artificial reefs, mangroves, estuaries and sandy shores.
Data collection. The PSP database was built from a compilation of the E. itajara observations by volunteer divers, encouraged to post sighting records, photographs and videos about the species into a PSP web-based platform (see www.merosdobrasil.org). The form requested the following information: diver name and contact; sighting date; diving site, habitat type (natural or artificial), depth, number of E. itajara sighted and estimated TL. Volunteers were contacted regarding doubts and the reliability of the information, and were requested to confirm the reported data. Incomplete or inaccurate questionnaires were discarded, as well as those with inconsistent information. Concomitantly, divers were encouraged to report sightings, posters were distributed to dive centers, and announcements were published in a specialized diving magazine and at research institutions in the States of Bahia, Pernambuco, and Santa Catarina stating the purpose of the study.
The sighting locations were inserted by divers on PSP form into a web map, through Google Maps tool, populated the map, providing sites and abundances of E. itajara at natural and artificial habitats.
Data analysis. Sites were categorized as artificial or natural habitats. Artificial habitat was defined as man-made structure, and included shipwrecks, concrete reefs and oil & gas structures. Size classes were analyzed through categories according TL, juvenile (<50 cm), subadult (50 - 100 cm), adult (101 - 150 cm) and large adult (151 - 200 cm). We used these classes interval because estimates tended to be informed at intervals of 50 cm (ex. 100, 150 cm). Depths were categorized into three ranges: shallow (0 - 14 m), mid depth (15 - 29 m), deep (≥ 30 m). For classification purposes, we considered herein an aggregation as the occurrence of more than five individuals at the same location. Goliath grouper TL between natural and artificial habitats was modeled as a function of the explanatory variable depth using generalized additive models (GAMs), performed with gam function from the mgcv package (Wood, 2006Wood, S. N. 2006. Generalized additive models: an introduction with R. Boca Raton, FL: Chapmann & Hall.) in software R (R Core Development Team, 2013). GAMs are semi parametric models where the dependent variable is linked to an additive predictor through a nonlinear link function (Dalla Rosa et al., 2012Dalla Rosa, L., J. K. B. Ford & A. W. Trites. 2012. Distribution and relative abundance of humpback whales in relation to environmental variables in coastal British Columbia and adjacent waters. Continental Shelf Research, 36: 89-104.). To evaluate changes in abundance between habitats, differences among depth categories were tested using non-parametric ANOVA (Kruskal-Wallis test). Tests were performed at a significance level of p < 0.05.
Results
Between 2005 and 2011, 188 reports from 84 volunteers were collected with a total of 306 individuals of E. itajara observed at 77 different sites along 13 Brazilian states. Reports were provided by recreational (n = 142), scientific (n = 31) and commercial divers (n = 15). Because reports with the presence of E. itajara are sporadic, it was not possible to obtain information about sampling efforts. The highest numbers of reports, 56%, came from the Northeast Brazil, followed by South, 23% and Southeast, 21%. Fernando de Noronha and Abrolhos, both National Marine Parks (NMPs) in Northeast, sheltered the majority of sites, with 17% and 14% respectively (Fig. 1).


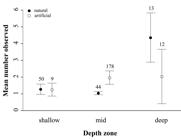
The maximum depth of species occurence was 38 m. Overall, goliath grouper were sighted 63% (n = 222) in shallow water (0 - 14 m). Individuals sighted in artificial habitats were at an average depth of 21.5 m (±5.1 s.d.), while in natural reefs, 16.3 m (±9.6 s.d.) (Fig. 3). There is a significant asymptotic relationship between TL and depth at natural habitats (F = 22.24, p < 0.0001; Fig. 4a). On the other hand, a clear nonlinear and non-significant relationship was observed at artificial habitats (F = 1.58; p = 0.18; Fig. 4b). Large adults were deeper, at average depth of 22.3 m (±6.7 s.d.), while juveniles and subadults were observed at an average depth of 18 m (±7.48 s.d.) (Fig. 4c) with no significant differences.
Observed E. itajara ranged from 10 to 200 cm TL and the most commonly reported sizes were subadults (60%) and adults (28%), with large adults (10%) and juveniles (2%) less represented. The highest number of individuals in an aggregation over artificial habitats was 10 and mean TL for all artificial habitat aggregations was 115.7 cm (±37.3 s.d.). For natural habitats, subadult and adult were the most frequent size classes observed (46% and 42%, respectively) (Fig. 4d). Subadults represents 65% of sightings at artificial habitat. The highest number of aggregated individuals at natural habitats was 11, at Gajiru rocky reefs, State of Sergipe. On natural habitats, average TL of aggregations was 110.6 cm (±39.8 s.d.), with 67% represented by subadult and 20% adult.
Overall, mean number of individuals observed was significantly higher at artificial habitat (mean = 1.93) than natural (mean = 1.27) (X2 = 8.08; p < 0.0001; Fig. 4d). The abundance between habitat types in shallow and deep strata was not significantly different (p > 0.05), but it was for the mid depth strata (X2 = 7.53; p < 0.0001; Fig. 5).
In general, sightings of a single individual were the most common situation (n = 146; 77%), followed by two to five specimens (n = 35; 18%) and seven aggregations accounted for six to eleven individuals, six of the ten smallest individuals (juveniles) were reported from estuaries.
At the monobuoy (a floating platform anchored offshore where tankers moor to unload oil) in south Brazil, aggregations were sighted in January 2009. At 'Parque dos Meros' artificial reefs, aggregations were registered in September 2008, June 2009, January 2010, and February 2010. Regarding natural reefs, a single record of an aggregation occurred at the State of Sergipe in January 2005, with 11 individuals.


Discussion
Our results revealed that habitat type influenced the relationship between TL and depth of occurrence of E. itajara. For artificial habitats there was a non-significant nonlinear relationship between TL and depth. The presence of artificial structures influences habitat choice, and the association of this species with artificial habitats is widely described in the literature (Carvalho-Filho, 1999Carvalho-Filho, A. 1999. Peixes: Costa brasileira. 3ª ed. São Paulo, Editora Melro.; Eklund & Schull, 2001Eklund, A. M. & J. Schull. 2001. A stepwise approach to investigate the movement patterns and habitat utilization of goliath grouper, Epinephelus itajara, using conventional tagging, acoustic telemetry and satellite tracking. Pp. 189-216. In: Nielsen, J. L. (Ed.). Electronic tagging and tracking in marine fisheries. Amsterdam, Kluwer Academic Publishers.; Félix-Hackradt & Hackradt, 2008Félix-Hackradt, F. C. & C. W. Hackradt. 2008. Populational study and monitoring of the goliath grouper, Epinephelus itajara (Lichtenstein, 1822), in the coast of Paraná, Brazil. Natureza & Conservação, 6: 31-46.). Artificial reefs are usually high relief reefs (Collins & Barbieri, 2010Collins, A. B. & L. R. Barbieri. 2010. Behavior, habitat, and abundance of the Goliath Grouper, Epinephelus itajara, in the Central Eastern Gulf of Mexico. Fish and Wildlife Research Institute, Florida Fish & Wildlife Conservation Commission.; Koenig et al., 2011Koenig, C. C., F. C. Coleman & K. Kingon. 2011. Pattern of recovery of the goliath grouper Epinephelus itajara population in the southeastern US. Bulletin of Marine Science, 87: 891-911.), and this vertical structure seems to exert an attraction to the species. The habitats reported in the present study (concrete reefs, shipwrecks and oil-gas structures) consisted of structures which can provide substantial shelter, especially for larger individuals, which demand habitats with abundant holes and caves sufficiently large to accommodate adult-sized individuals (Koenig & Coleman, 2009Koenig, C. C. & F. C. Coleman. 2009. Population density, demographics, and predation effects of adult goliath grouper. National Oceanic and Atmospheric Administration, Marfin Project Final Report.). This is not necessarily available in shallow reefs.
Abundances of E. itajara by sighting at artificial habitats were significantly higher than at natural ones, especially in mid depths. Such correlation between artificial habitat and the abundance of E. itajara was also observed in the central eastern Gulf of Mexico (Collins, 2008Collins, A. B. 2008. A preliminary assessment of the abundance and size distribution of goliath grouper Epinephelus itajara within a defined region of the Central Eastern Gulf of Mexico. Proceedings of the 61st Gulf and Caribbean Fisheries Institute.). In contrast, Koenig et al. (2011)Koenig, C. C., F. C. Coleman & K. Kingon. 2011. Pattern of recovery of the goliath grouper Epinephelus itajara population in the southeastern US. Bulletin of Marine Science, 87: 891-911. observed that adults prefer high-relief habitats that provide shelter, but observed no preference between the high-relief artificial and high-relief natural reefs.
Although 'Parque dos Meros' and Fernando de Noronha accounted for the largest number of reports (25 and 24; see Fig. 1), it does not necessarily mean that the species is most abundant at these points. Such sites are frequently visited by recreational scuba divers, mainly during austral summer. Based in high site fidelity for E. itajara (Koenig et al., 2011Koenig, C. C., F. C. Coleman & K. Kingon. 2011. Pattern of recovery of the goliath grouper Epinephelus itajara population in the southeastern US. Bulletin of Marine Science, 87: 891-911.) and the largest amount of reports on sites that diving tourism exists, it is possible that reports include several sightings of the same fish. Resightings of individual E. itajara at specific sites have been observed through the years, and images provide the photo-identification of individuals through scars (Giglio et al., 2014Giglio, V. J., J. Adelir-Alves & A. A. Bertoncini. 2014. Using scars to photo-identify the goliath grouper, Epinephelus itajara. Marine Biodiversity Records, 7, e108. DOI: 10.1017/S1755267214001080.
https://doi.org/10.1017/S175526721400108...
), showing high degrees (> 4 years) of residence.
Whilst size at first maturity has not been reported for the South Atlantic stock, Bullock et al. (1992)Bullock, L. H., M. D. Murphy, M. F. Godcharles & M. E. Mitchell. 1992. Age, growth and reproduction of jewfish Epinephelus itajara in the eastern Gulf of Mexico. Fisheries Bulletin, 90: 243-249. found that individuals around 115.5 cm were observed to be sexually mature in the Eastern Gulf of Mexico. During this study, the majority of observed fish were medium-sized (100 to 150 cm TL), suggesting that most fish documented were adults. Large adults were observed in all depth ranges (sightings were reported from 5 to 30 m). Juveniles are often found in estuarine areas (Eklund & Schull, 2001Eklund, A. M. & J. Schull. 2001. A stepwise approach to investigate the movement patterns and habitat utilization of goliath grouper, Epinephelus itajara, using conventional tagging, acoustic telemetry and satellite tracking. Pp. 189-216. In: Nielsen, J. L. (Ed.). Electronic tagging and tracking in marine fisheries. Amsterdam, Kluwer Academic Publishers.; Frias-Torres, 2006Frias-Torres, S. 2006. Habitat use of juvenile goliath grouper Epinephelus itajara in the Florida Keys, USA. Endangered Species Research, 2: 1-6.). However, ontogenetic offshore migration is not necessarily related to growth. For juveniles, previous studies observed individuals out of estuaries at wide range depths (Collins, 2008Collins, A. B. 2008. A preliminary assessment of the abundance and size distribution of goliath grouper Epinephelus itajara within a defined region of the Central Eastern Gulf of Mexico. Proceedings of the 61st Gulf and Caribbean Fisheries Institute.; Koenig et al., 2011Koenig, C. C., F. C. Coleman & K. Kingon. 2011. Pattern of recovery of the goliath grouper Epinephelus itajara population in the southeastern US. Bulletin of Marine Science, 87: 891-911.), indicating that E. itajara may leave nursery habitats earlier than previously believed (100 cm TL; Koenig et al., 2007Koenig, C. C., F. C. Coleman, A. M. Eklund, J. Schull & J. Ueland. 2007. Mangroves as essential nursery habitat for goliath grouper (Epinephelus itajara), Bulletin of Marine Science, 80: 567-586.).
Goliath grouper aggregations were observed in three sites during austral summer, spring and winter. The species aggregates during spawning season, which is suspected to take place during the summer months within the study region. In South Brazil, at Santa Catarina State and 'Parque dos Meros' artificial reefs, aggregations were observed during austral summer and winter. However, our results have limitations once sampling effort is not available by seasons.
In Brazil, small aggregations have been reported by artisanal fishermen as reproductive events during austral summer (December - March; Reuss-Strenzel & Assunção, 2008Reuss-Strenzel, G. M. & M. F. Assunção. 2008. Etnoconhecimento ecológico dos caçadores submarinos de Ilhéus, Bahia, como subsídio à preservação do mero (Epinephelus itajara Lichtenstein, 1822). Revista da Gestão Costeira Integrada, 8: 203-219.; Gerhardinger et al., 2009Gerhardinger, L. C., M. Hostim-Silva, R. P. Medeiros, J. Matarezi, A. A. Bertoncini, M. O. Freitas & B. P. Ferrreira. 2009. Fishers resource mapping and goliath grouper Epinephelus itajara (Serranidae) conservation in Brazil. Neotropical Ichthyology, 7: 93-102.). However, aggregations in Brazil have also been observed during autumn (Félix-Hackradt & Hackradt, 2008Félix-Hackradt, F. C. & C. W. Hackradt. 2008. Populational study and monitoring of the goliath grouper, Epinephelus itajara (Lichtenstein, 1822), in the coast of Paraná, Brazil. Natureza & Conservação, 6: 31-46.) and spring (J. Adelir-Alves, pers. comm.), demanding more specific investigations on the nature of these aggregations. These sites may shelter small resident aggregations throughout the year, possibly due to the availability of shelter and food resources, besides being close to estuaries.
Mapping spawning aggregation sites and seasons will be of importance when formulating conservation strategies to curb illegal fishing practices. Given that illegal fishing of aggregations are regularly observed in Brazil at sites still unknown to science (IBAMA, 2009IBAMA. 2009. Ibama apreende seis toneladas de peixe no Maranhão, IBAMA, Brasília. Available from: http://www.ibama.gov.br/noticias-2009/ibama-apreende-seis-toneladas-de-peixe-no-maranhao (8 Jun 2013).
http://www.ibama.gov.br/noticias-2009/ib...
; ICMBio, 2011ICMBio. 2011. Parceria proporciona apreensão de quatro toneladas de mero. Boletim interno do ICMBio, 135: 3.; authors pers. obs.), data on species occurrence, seasonal distribution, and site preferences will provide the necessary information to better apply surveillance efforts and other E. itajara conservation initiatives.
Regarding quality and validity of the data collected by volunteers, a series of features in our study led us to conclude that the data presented here are reliable. Firstly, the distinctive morphological characteristics and behavior reduces the probability of E. itajara misidentifications. Secondly, the requested information such as dive site, depth, and habitat are details most divers habitually record in personal dive logs. Finally, we did not observe discrepancies regarding TL between the information of researchers and recreational divers. However, the main limitation of our study is that volunteer diver surveys are often concentrated in areas of prime habitat, are more frequent during certain seasons, and so being rarely distributed evenly through space and time.
This study provides the first broad spatial scale visualization of habitat use and abundance of E. itajara along the Brazilian coastline. Our results underscore the importance of developing conservation measures addressing specific habitats and locations to boost population recovery. Artificial structures might be strategic habitats for goliath grouper conservation policies, but should be properly protected from illegal fishing practices and habitat degradation as a fundamental strategy for the recovery of these populations, i.e., by spatially bounded and appropriately enforced governing regimes. Otherwise, artificial reefs will act as unintentional fishing enhancement devices. We believe this concern should be entirely and comprehensively thought as part of the step-zero process of environmental licensing for artificial reefs. As Gerhardinger et al. (2009)Gerhardinger, L. C., R. Medeiros., R. C. Marenzi., A. A. Bertoncini & M. Hostim-Silva. 2006. Local Ecological Knowledge on the Goliath Grouper Epinephelus itajara (Teleostei: Serranidae) in Southern Brazil. Neotropical Ichthyology, 4: 441-450. have suggested the 'Parque dos Meros' artificial reefs are of high conservation concern, as large goliath groupers attracted to these artificial reefs have become easy targets for spearfishermen. Nonetheless, the occurrence of this docile, large-sized and emblematic fish adds an important attraction to any diving sites. Therefore, only if they are properly governed, goliath grouper abundant seascapes could become an economic alternative allied with conservation outcomes.
We thank the PSP collaborators, especially I. Cerutti, A. Calomeno, F. Freitas, S. Amarante, and E. Marcovaldi. Environmental monitors and dive staff of Abrolhos National Marine Park for reports. The Ecomar NGO for logistical support. The research was granted financial support from Petrobras and Fundação Grupo o Boticário de Proteção à Natureza (project 0774_20081). We also thank M. Cantor, O. J. Luiz and M. Borgonha for their comments, as well as two anonymous referees that greatly improved the manuscript.
Literature Cited
- Amaral, A. C. Z. & S. Jablonski. 2005. Conservation of Marine and Coastal Biodiversity in Brazil. Conservation Biology, 19: 625-631.
- Auster, P. J., B. X. Semmens & K. Barber. 2005. Pattern in the co-occurrence of fishes inhabiting the coral reefs of Bonaire, Netherlands Antilles. Environmental Biology of Fishes, 74: 187-194.
- Bullock, L. H., M. D. Murphy, M. F. Godcharles & M. E. Mitchell. 1992. Age, growth and reproduction of jewfish Epinephelus itajara in the eastern Gulf of Mexico. Fisheries Bulletin, 90: 243-249.
- Carlson, J. K., S. J. B. Gulak, C. A. Simpfendorfer, R. D. Grubbs & J. G. Romine. 2013. Movement patterns and habitat use of smalltooth sawfish, Pristis pectinata, determined using pop-up satellite archival tags. Aquatic Conservation: Marine and Freshwater Ecosystems, DOI: 10.1002/aqc.2382.
» https://doi.org/10.1002/aqc.2382 - Carvalho-Filho, A. 1999. Peixes: Costa brasileira. 3ª ed. São Paulo, Editora Melro.
- Collins, A. B. 2008. A preliminary assessment of the abundance and size distribution of goliath grouper Epinephelus itajara within a defined region of the Central Eastern Gulf of Mexico. Proceedings of the 61st Gulf and Caribbean Fisheries Institute.
- Collins, A. B. & L. R. Barbieri. 2010. Behavior, habitat, and abundance of the Goliath Grouper, Epinephelus itajara, in the Central Eastern Gulf of Mexico. Fish and Wildlife Research Institute, Florida Fish & Wildlife Conservation Commission.
- Craig, M. T., R. T. Graham, R. A. Torres, J. R. Hyde, M. O. Freitas, B. P. Ferreira, M. Hostim-Silva, L. C. Gerhardinger, A. A. Bertoncini & D. R. Robertson. 2009. How many species of goliath grouper are there? Cryptic genetic divergence in a threatened marine fish and the resurrection of a geopolitical species. Endangered Species Research, 7: 167-174.
- Dalla Rosa, L., J. K. B. Ford & A. W. Trites. 2012. Distribution and relative abundance of humpback whales in relation to environmental variables in coastal British Columbia and adjacent waters. Continental Shelf Research, 36: 89-104.
- Darwal, W. R. T. & N. T. Dulvy. 1996. An evaluation of the suitability of non-specialist volunteer researchers for coral reef fish surveys. Mafia Island, Tanzania - A case study. Biological Conservation, 78: 223-231.
- Eklund, A. M. & J. Schull. 2001. A stepwise approach to investigate the movement patterns and habitat utilization of goliath grouper, Epinephelus itajara, using conventional tagging, acoustic telemetry and satellite tracking. Pp. 189-216. In: Nielsen, J. L. (Ed.). Electronic tagging and tracking in marine fisheries. Amsterdam, Kluwer Academic Publishers.
- Félix-Hackradt, F. C. & C. W. Hackradt. 2008. Populational study and monitoring of the goliath grouper, Epinephelus itajara (Lichtenstein, 1822), in the coast of Paraná, Brazil. Natureza & Conservação, 6: 31-46.
- Foster - Smith, J. & S. M. Evans. 2003. The value of marine ecological data collected by volunteers. Biological Conservation, 113: 199-213.
- Frias-Torres, S. 2006. Habitat use of juvenile goliath grouper Epinephelus itajara in the Florida Keys, USA. Endangered Species Research, 2: 1-6.
- Frias-Torres, S. 2012. Should the Critically Endangered Goliath grouper Epinephelus itajara be culled in Florida? Oryx, 1: 1-8.
- Gerhardinger, L. C., M. Hostim-Silva, R. P. Medeiros, J. Matarezi, A. A. Bertoncini, M. O. Freitas & B. P. Ferrreira. 2009. Fishers resource mapping and goliath grouper Epinephelus itajara (Serranidae) conservation in Brazil. Neotropical Ichthyology, 7: 93-102.
- Gerhardinger, L. C., R. Medeiros., R. C. Marenzi., A. A. Bertoncini & M. Hostim-Silva. 2006. Local Ecological Knowledge on the Goliath Grouper Epinephelus itajara (Teleostei: Serranidae) in Southern Brazil. Neotropical Ichthyology, 4: 441-450.
- Giglio, V. J., J. Adelir-Alves & A. A. Bertoncini. 2014. Using scars to photo-identify the goliath grouper, Epinephelus itajara. Marine Biodiversity Records, 7, e108. DOI: 10.1017/S1755267214001080.
» https://doi.org/10.1017/S1755267214001080 - Goffredo, S., F. Pensa, P. Neri, A. Orlandi, M. S. Gagliardi, A. Velardi, C. Piccinetti & F. Zaccanti. 2010. Unite research with what citizens do for fun: ''recreational monitoring'' of marine biodiversity. Ecological Applications, 20: 2170-2187.
- Goffredo, A., C. Piccinetti & F. Zaccanti. 2004. Volunteers in marine conservation monitoring: A study of the distribution of seahorses carried out in collaboration with recreational scuba divers. Conservation Biology, 18: 1492-1503.
- Heemstra, P. C. & J. E. Randall. 1993. FAO species catalogue: Groupers of the world (Family Serranidae, subfamily Epinephelinae). An annotated and illustrated catalogue of the grouper, rockcod, hind, coral grouper and lyretail species known to date. FAO Fisheries Synopis, 16: 1-382.
- Holt, B. G., R. Rioja-Nieto, M. A. MacNeil, J. Lupton & C. Rahbek. 2013. Comparing diversity data collected using a protocol designed for volunteers with results from a professional alternative. Methods in Ecology and Evolution, 4: 383-392.
- Hostim-Silva M., A. A. Bertoncini, L. C. Gerhardinger & L. F. Machado. 2005. The 'Lord of the Rock's' conservation program in Brazil: the need for a new perception of marine fishes. Coral Reefs 24, 7.
- Hussey, N. E., N. Stroh, R. Klaus, T. Chekchak & S. T. Kessel. 2013. SCUBA diver observations and placard tags to monitor grey reef sharks, Carcharhinus amblyrhynchos, at Sha'ab Rumi, The Sudan: assessment and future directions. Journal of the Marine Biological Association of the United Kingdom, 93, 299-308.
- IBAMA. 2009. Ibama apreende seis toneladas de peixe no Maranhão, IBAMA, Brasília. Available from: http://www.ibama.gov.br/noticias-2009/ibama-apreende-seis-toneladas-de-peixe-no-maranhao (8 Jun 2013).
» http://www.ibama.gov.br/noticias-2009/ibama-apreende-seis-toneladas-de-peixe-no-maranhao - ICMBio. 2011. Parceria proporciona apreensão de quatro toneladas de mero. Boletim interno do ICMBio, 135: 3.
- IUCN. 2013. The IUCN Red List of Threatened Species. International Union for Conservation of Nature and Natural Resources. Available from: http://www.iucnredlist.org/details/195409/0 (10 Aug 2013).
» http://www.iucnredlist.org/details/195409/0 - Koenig, C. C. & F. C. Coleman. 2009. Population density, demographics, and predation effects of adult goliath grouper. National Oceanic and Atmospheric Administration, Marfin Project Final Report.
- Koenig, C. C., F. C. Coleman, A. M. Eklund, J. Schull & J. Ueland. 2007. Mangroves as essential nursery habitat for goliath grouper (Epinephelus itajara), Bulletin of Marine Science, 80: 567-586.
- Koenig, C. C., F. C. Coleman & K. Kingon. 2011. Pattern of recovery of the goliath grouper Epinephelus itajara population in the southeastern US. Bulletin of Marine Science, 87: 891-911.
- Luiz, O. J., A. P. Balboni, G. Kodja, M. Andrade & H. Marum. 2009. Seasonal occurrences of Manta birostris (Chondrichthyes: Mobulidae) in southeastern Brazil. Ichthyological Research, 56: 96-99.
- McClenachan, L. 2009. Historical declines of goliath grouper populations in South Florida, USA. Endangered Species Research, 7: 175-181.
- Pina-Amargós, F. & G. González-Sansón. 2009. Movement of goliath grouper (Epinephelus itajara) in southeast Cuba: implications for its conservation. Endangered Species Research, 7: 243-247.
- Reuss-Strenzel, G. M. & M. F. Assunção. 2008. Etnoconhecimento ecológico dos caçadores submarinos de Ilhéus, Bahia, como subsídio à preservação do mero (Epinephelus itajara Lichtenstein, 1822). Revista da Gestão Costeira Integrada, 8: 203-219.
- Ruttenberg, B. I., P. J. Schofield, J. L. Akins, A. Acosta, M. W. Feeley, J. Blondeau, S. G. Smith & J. S. Ault. 2012. Rapid invasion of Indo-Pacific lionfishes (Pterois volitans and Pterois miles) in the Florida Keys, USA: evidence from multiple pre-and post-invasion data sets. Bulletin of Marine Science, 88: 1051-1059.
- Sadovy, Y. & A. Eklund. 1999. Synopsis of biological data on the Nassau Grouper, Ephinephelus striatus, and the Jewfish, E. itajara. NOAA Technical Report.
- Stallings, C. 2009. Fishery-independent data reveal negative effect of human population density on caribbean predatory fish communities. PLoS ONE, 4: e5333.
- Thompson, W. L . 2004. Sampling rare or elusive species. Island Press: Washington, D.C.
- Ward-Paige, C. A., C. Mora, H. K. Lotze, C. Pattengill-Semmens, L. McClenachan, E. Arias-Castro & R. A. Myers. 2010. Large-Scale Absence of Sharks on Reefs in the Greater- Caribbean: A Footprint of Human Pressures. PLoS ONE, 5: e11968.
- Wiley, T. & C. Simpfendorfer. 2010. Using public encounter data to direct recovery efforts for the endangered smalltooth sawfish Pristis pectinata. Endangered Species Research, 12: 179-191.
- Wood, S. N. 2006. Generalized additive models: an introduction with R. Boca Raton, FL: Chapmann & Hall.
Publication Dates
-
Publication in this collection
Oct-Dec 2014
History
-
Received
24 Sept 2013 -
Accepted
29 July 2014





