Abstracts
TgROP2 is an intracellular protein associated with rhoptries of Toxoplama gondii and an antigen component of a candidate vaccine for toxoplasmosis. The purpose of the present study was to evaluate the efficacy of rTgROP2 to stimulate humoral and cellular immune responses in BALB/c mice via intranasal injection. TgROP2 partial coding sequence was (196-561) amplified by PCR from genomic T. gondii RH strain DNA and cloned into the pTrcHis expression vector. Escherichia coli Rosetta 2 cells transformed with pTrcHis-TgROP2 showed high levels (~1 mg.mL-1) of recombinant protein after 4 hours of IPTG induction. Recombinant TgROP2 exhibited an apparent Mr equal to 54 kDa. In order to test immunogenicity of the recombinant protein, 10 BALB/c mice received 10 µg of rROP2 protein + 10 µg of Quil-A via intranasal injection. Doses were administered at days 0, 21, and 42. Three animals were euthanized and used to evaluate cell-ular immune response on day 62. Five (50%) and two (20%) out of ten animals produced IgG (DO mean = 0.307; cut-off = 0.240) and IgA (DO mean = 0.133, cut-off = 0.101), respectively, by ELISA on day 62. The proliferation of splenocytes revealed high stimulation index (SI) when co-cultured with 5, 10 and 15 µg.mL-1 of rTgROP2. These results indicate that intranasal immunization with recombinant protein ROP2 plus Quil-A can elicit both cellular and humoral immune responses in BALB/c mice.
Cloning; expression; recombinant protein; rhoptry; rROP2
TgROP2 é uma proteína localizada nas roptrias do Toxoplasma gondii, sendo um antígeno candidato a componente de uma vacina contra a toxoplasmose. O objetivo do presente estudo foi avaliar a eficácia da TgROP2 recombinante em estimular a resposta imune celular e humoral de camundongos BALB/c após estímulo intranasal. A sequência da TgROP2 foi amplificada pela PCR a partir da cepa RH e clonada em vetor de expressão pTrc-His. Após a transformação em Escherichia coli- Rosetta 2, a pTrcHis-TgROP2 exibiu alto nível de expressão após 4 horas de indução com IPTG. A proteína recombinante apresentou uma massa molecular aparente de aproximadamente 54 kDa. Para avaliar a imunogenicidade dessa proteína recombinante, 10 camundongos receberam, pela via intranasal, 10 µg da rROP2 associado a 10 µg de Quil-A. Três doses foram realizadas nos dias 0, 21 e 42. No dia 62 do experimento, três animais foram eutanasiados para avaliar as respostas imune celular e humoral. Cinco (50%) e dois (20%) dos 10 animais apresentaram níveis de IgG (DO média = 0,307; ponto de corte = 0,240) e IgA (DO média = 0,133; ponto de corte = 0,101) acima do ponto de corte no ELISA no dia 62. A proliferação de esplenócitos revelou altos Índices de Estimulação (SI), quando as células foram cultivadas com 5, 10 e 15 µg.mL-1 de rTgROP2. Os resultados obtidos indicam que a via nasal pode estimular tanto a resposta imune celular como a humoral.
Clonagem; expressão; proteína recombinante; roptrias; rROP2
FULL ARTICLE
Toxoplasma gondii: humoral and cellular immune response of BALB/c mice immunized via intranasal route with rTgROP2
Toxoplasma gondii: avaliação da resposta imune humoral e celular de camundongos BALB/c imunizados pela via nasal com rTgROP2
Michelle IgarashiI; Dauton Luiz ZulpoI; Ivo Alexandre Leme da CunhaI; Luiz Daniel BarrosI; Vanessa Figueredo PereiraI; Alessandra TarodaI; Italmar Teodorico NavarroI; Odilon VidottoI; Marilda Carlos VidottoI; Mark Christopher JenkinsII; João Luis GarciaI
ILaboratório de Protozoologia, Departamento de Medicina Veterinária Preventiva, Universidade Estadual de Londrina UEL
IIAnimal Parasitic Disease Laboratory, United States Department of Agriculture
Corresponding author Corresponding author: João Luis Garcia Laboratório de Protozoologia Departamento de Medicina Veterinária Preventiva Universidade de Londrina UEL CP 6001, CEP 86050-970 Londrina - PR, Brazil e-mail: jlgarcia@uel.br
ABSTRACT
TgROP2 is an intracellular protein associated with rhoptries of Toxoplama gondii and an antigen component of a candidate vaccine for toxoplasmosis. The purpose of the present study was to evaluate the efficacy of rTgROP2 to stimulate humoral and cellular immune responses in BALB/c mice via intranasal injection. TgROP2 partial coding sequence was (196-561) amplified by PCR from genomic T. gondii RH strain DNA and cloned into the pTrcHis expression vector. Escherichia coli Rosetta 2 cells transformed with pTrcHis-TgROP2 showed high levels (~1 mg.mL1) of recombinant protein after 4 hours of IPTG induction. Recombinant TgROP2 exhibited an apparent Mr equal to 54 kDa. In order to test immunogenicity of the recombinant protein, 10 BALB/c mice received 10 µg of rROP2 protein + 10 µg of Quil-A via intranasal injection. Doses were administered at days 0, 21, and 42. Three animals were euthanized and used to evaluate cell-ular immune response on day 62. Five (50%) and two (20%) out of ten animals produced IgG (DO mean = 0.307; cut-off = 0.240) and IgA (DO mean = 0.133, cut-off = 0.101), respectively, by ELISA on day 62. The proliferation of splenocytes revealed high stimulation index (SI) when co-cultured with 5, 10 and 15 µg.mL1 of rTgROP2. These results indicate that intranasal immunization with recombinant protein ROP2 plus Quil-A can elicit both cellular and humoral immune responses in BALB/c mice.
Keywords: Cloning, expression, recombinant protein, rhoptry, rROP2.
RESUMO
TgROP2 é uma proteína localizada nas roptrias do Toxoplasma gondii, sendo um antígeno candidato a componente de uma vacina contra a toxoplasmose. O objetivo do presente estudo foi avaliar a eficácia da TgROP2 recombinante em estimular a resposta imune celular e humoral de camundongos BALB/c após estímulo intranasal. A sequência da TgROP2 foi amplificada pela PCR a partir da cepa RH e clonada em vetor de expressão pTrc-His. Após a transformação em Escherichia coli- Rosetta 2, a pTrcHis-TgROP2 exibiu alto nível de expressão após 4 horas de indução com IPTG. A proteína recombinante apresentou uma massa molecular aparente de aproximadamente 54 kDa. Para avaliar a imunogenicidade dessa proteína recombinante, 10 camundongos receberam, pela via intranasal, 10 µg da rROP2 associado a 10 µg de Quil-A. Três doses foram realizadas nos dias 0, 21 e 42. No dia 62 do experimento, três animais foram eutanasiados para avaliar as respostas imune celular e humoral. Cinco (50%) e dois (20%) dos 10 animais apresentaram níveis de IgG (DO média = 0,307; ponto de corte = 0,240) e IgA (DO média = 0,133; ponto de corte = 0,101) acima do ponto de corte no ELISA no dia 62. A proliferação de esplenócitos revelou altos Índices de Estimulação (SI), quando as células foram cultivadas com 5, 10 e 15 µg.mL1 de rTgROP2. Os resultados obtidos indicam que a via nasal pode estimular tanto a resposta imune celular como a humoral.
Palavras-chave: Clonagem, expressão, proteína recombinante, roptrias, rROP2.
Introduction
There is considerable interest in developing a vaccine against Toxoplasma gondii infection due to the impact of toxoplasmosis on humans and animals. An effective vaccine should protect against both acute and chronic infection (HAUMONT et al., 2000). ROP2 antigen is expressed by T. gondii tachyzoites, bradyzoites, and sporozoites, and has been proposed as a vaccine candidate against toxoplasmosis (SAAVEDRA et al., 1996). The use of rTgROP2 antigen has protected mice against chronic T. gondii infection (IGARASHI et al., 2008b). This and other studies have shown that ROP2 vaccination leads to a decrease in tissue cysts burden (IGARASHI et al., 2008b; MARTIN et al., 2004). Therefore, rTgROP2 is a viable candidate for an effective vaccine for acute toxoplasmosis infection.
The natural site of T. gondii infection is the mucosal surface of the intestine wherein immunoglobulin A (IgA) specific for T. gondii is synthesized. Protective immunity acquired after natural infection with T. gondii points to the importance to develop a vaccine that can stimulate mucosal defenses (BOURGUIN et al., 1993). Oral vaccine delivery appears to be a common route of antigen presentation, but it requires administration of massive amounts of antigen (DEBARD et al., 1996). The intranasal route is an interesting alternative vaccination route and is more effective than intragastric immunization in that it generates an earlier and stronger mucosal immune response (HIRABAYASHI et al., 1990; WU; RUSSEL, 1997). The intranasal route requires a smaller amount of antigen than the oral one; it promotes both systemic and mucosal immune responses to an antigen (WU; RUSSEL, 1997) and prevents the development of oral tolerance. Primary T. gondii infection induces a protective immune response in BALB/c mice, which prevents transplacental transmission of parasite to the fetus during subsequent reinfection, making it a particularly relevant model of human infection (ROBERTS; ALEXANDER, 1992).
Smith et al. (1998) have shown that oral administration of free Quil-A together with OVA reproduced most of local and systemic immune responses obtained with ISCOM and OVA. The saponin adjuvant Quil-A is obtained from the bark of a tree Quillaja saponaria. Quil-A is a widely used veterinary adjuvant, low cost, simple to formulate and generally safe (COX; COULTER, 1997). Garcia et al. (2007) used crude T. gondii rhoptry proteins plus Quil-A via intranasal route to evaluate the control of oocyst shed in domestic cats. Two out of three cats did not shed oocysts after being immunized with three doses of the vaccine, and vaccinated cats had an estimated protection of 65%.
In the present study we evaluated humoral and cellular immune responses in BALB/c mice immunized via intranasal route with rROP2
Material and Methods
1. Toxoplasma gondii strains
Toxoplasma gondii RH strain tachyzoites were obtained from peritoneal fluid of infected Swiss Webster mice. Tachyzoites were passed three times through a 26 gauge needle, and washed twice with 10 mM phosphate buffered saline (PBS, pH 7.5) by centrifugation. The pellet was suspended and washed twice in PBS; after washing, tachyzoites were diluted to 105 tachyzoites.mL1 prior to DNA extraction. ME-49 strain of T. gondii was used to challenge mice with tissue brain cysts.
2. Construction of plasmids
The DNA sequence of the gene encoding T. gondii ROP2 rhoptry antigen was obtained from Genbank database (Accession number: Z36906). It was amplified using rop2 forward (5'ATCGAATTCACGGATCCTGGAGAC3'-introduced EcoRI recognition site, underlined) and reverse (5'TGAAAGCTTTCATGCCGGTTCTCC3' introduced HindIII recognition site, underlined) primers in a standard PCR amplification. Each PCR mixture (25 µL) consisted of 10 mM Tris-HCl pH 8.3, 50 mM MgCl2, 0.5 µM of each primer, 0.2 mM nucleoside triphosphates (d-NTPs), and 1.25 units of Taq DNA polymerase (Invitrogen, Life Technologies, USA). PCR conditions were 3 minutes denaturation at 94 ºC, followed by 30 cycles of 30 seconds each at 94 ºC, 30 seconds at 55 ºC, 1 minute at 72 ºC; followed by a final 5 minutes extension at 72 ºC. After amplification, 5 µL aliquots from each reaction were analyzed by eletrophoresis in 7.5% polyacrylamide mini-gels. DNA fragments were stained with ethidium bromide solution and visualized under UV light (254 nm) (Biophotonics, Inc., Ann Arbor, Michigan). The PCR product was obtained with 1103 pb size. This fragment was digested overnight with EcoRI and HindIII endonucleases and ligated into pTrcHis B (Invitrogen, Life Technologies, USA) following the manufacturer's recommendations. PCR products were purified from amplification reactions using a QIAquick PCR purification kit (Qiagen, QIAGEN Biotecnologia Brasil Ltda., São Paulo, Brazil) or from agarose gels after restriction enzyme digestion and gel electrophoresis using a QIAquick DNA gel purification kit (Qiagen, QIAGEN Biotecnologia Brasil Ltda., São Paulo, Brazil). Purified PCR products were precipitated with 2 volumes of 100% ethanol and 0.1 volume of 3 M sodium acetate, washed with 70% ethanol, dried, and then suspended in 10 µL sterile water. Restriction enzyme digested PCR products were cloned into pTrcHis using T4 DNA ligase (Invitrogen, Life Technologies, USA), and introduced into Escherichia coli DH5 cells using standard transformation procedures (HANAHAN, 1983). E. coli DH5-α transformants harboring recombinant pTrcHis-TgROP2 plasmid were identified by colony-PCR using pTrcHis forward and reverse primers (flanking the insertion site). Plasmid DNA from positive clones was prepared from E. coli transformants using a DNA mini-plasmid kit (Qiagen, QIAGEN Biotecnologia Brasil Ltda., São Paulo, Brazil), and subjected to dideoxy sequencing using primers specific for pTrcHis vector. Maintenance of the open reading frame was confirmed by sequence analysis using Sequencher software prior to expression studies.
3. Culture conditions
Recombinant pTrcHis-TgROP2 plasmid DNA was transformed into E. coli Rosetta 2 cells. E. coli strain Rosetta cells transformed with pTrcHis/ROP2 were grown with vigorous shaking at 37 ºC in 50 mL LB supplemented with 100 µg.mL1 ampicillin and 100 µg.mL1 chloramphenicol to an OD600 = 0.8 and then induced with 1 mM (final concentration) isopropyl-Dthiogalactopyranoside (IPTG) for an additional 4 hours at 37 ºC. The cells were harvested by centrifugation at 2500 g for 5 minutes, and the pellets were suspended in 20 mM sodium phosphate and 500 mM sodium chloride pH 7.8, and lysed by 3 freezing-thawing cycles between a dry ice-ethanol bath and a 37 ºC water bath. The protein extraction was treated for 30 minutes at 25 ºC with 1 µg.mL1 DNase and RNase to destroy nucleic acid, and then pelleted by centrifugation at 3000 g for 15 minutes at 4 ºC. The soluble fraction after centrifugation was collected and used in subsequent NiNTA purification to isolate recombinant TgROP2 protein.
4. NiNTA purification of TgROP2
The soluble fraction was directly applied to NiNTA Superflow resin (Qiagen, QIAGEN Biotecnologia Brasil Ltda., São Paulo, Brazil) pre-equilibrated with 20 mM sodium phosphate, 500 mM sodium chloride, pH 7.8 (native binding buffer, NBB) for soluble samples, and allowed to bind by placing on a rocker for 30 minutes at room temperature. Unbound E. coli protein was removed from the column by gravity flow, and NiNTA agarose beads were washed with 5 column volumes of NBB, followed by 5 column volumes of 20 mM sodium phosphate, 500 mM sodium chloride, pH 6.8. The recombinant soluble antigen was eluted from resin by gravity flow with native elution buffer (200 mM monobasic sodium phosphate and 5 M NaCl pH 4.0).
5. Electrophoresis and western blot analysis
Recombinant TgROP2 antigen was subjected to SDS-polyacrylamide gel electrophoresis and colloidal Comassie staining following standard procedures. Protein concentrations were determined by BCA Protein Assay Reagent (Pierce Chemical Co.). After electrophoresis proteins were transferred to an Immobilon membrane (Millipore) on a semi-dry blotter apparatus (Invitrogen, Life Technologies, USA). The membrane was saturated with 5% non-fat dry milk (NFDM) in PBS for 30 minutes and then washed three times in PBS-Tween20 (PBS-Tw). The membrane was incubated for 1 hour with mouse anti-His tag antibody (Invitrogen, Life Technologies, USA) diluted in PBS-Tw solution (1:1000), followed by a 1 hour incubation with biotinylated anti-mouse IgG (Sigma Chemical Co.), followed by a 1 hour incubation with avidin peroxidase diluted 1:5000 in PBS-Tw (Sigma). All washes between incubation steps consisted of 5 minutes with PBS-Tw. Reactive bands were visualized by incubation of the blots with 0.06% 4-chloro-napthol and 0.02% hydrogen peroxide in PBS.
6. Experimental design
The present study was approved by Animal Ethics Committee at Universidade Estadual de Londrina (CEEA 47/06). Ten females BALB/c of 10 weeks of age were used in the experiment. The animals received via intranasal route 10 µg of recombinant ROP2 plus 10 µg of Quil-A as adjuvant. Each dose of immunogen was instilled into their nostrils with a micropipettor (6 µL.nostril1). Mice were immunized on days 0, 21 and 42. Their blood was collected by retro-orbital plexus puncture on days 0, 20, 42 and 62 and sera were stored at 20 ºC until used for analysis. On day 62 three mice were euthanized for cellular immune response evaluation.
7. Measurement of antibody response enzyme linked immunosorbent assay
ELISA was performed as previously described (GARCIA et al., 2006). Optimal dilutions were established using checkerboard titrations with dilutions of sera, antigen, and conjugates. rROP2 was used as an antigen to coat wells (5 µg.mL1) of microtiter plates. Flat-bottomed polystyrene 96-well microtitration plates (Nunc-Immuno Plate, MaxiSorp, Denmark) were coated with 0.1 mL of antigens (5 µg.mL1) diluted in 0.1 M carbonate buffer (pH 9.6) and incubated overnight at 4 ºC. The plates were washed three times with TBS-tween 20 (50 mM tris, 150 mM sodium chloride, and 0.05% tween 20, pH 7.4) and non-specific immune sites were blocked with buffer containing 8% non-fat dry milk. Control and test sera were diluted at 1:100 and 1:4 for IgG and IgA, respectively, in PBS-tween 20 plus 5% non-fat dry milk and then added to microtiter plates in duplicates, 0.1 mL in each well, and incubated for 1 hour at 37 ºC. After washing, anti-mouse IgG and IgA antibodies were diluted at 1:5000 in PBS-tween 20 plus 5% non-fat dry milk, and 0.1 mL was added to each well and incubated for 1 hour at 37 ºC. After washing, peroxidase activity was revealed by adding 0.1 mL of ortho-phenilenediamine solution (40 mg ortho-phenilenediamine/100 mL of 0.1 M phosphate citrate buffer, pH 6.0 and 40 µL of H2O2), and the reaction was blocked by adding 0.05 mL of 1 N HCl. Optical density (OD) was read at 490 nm in an ELISA microplate reader. Mean absorbance values were estimated and OD value was calculated as previously described (GARCIA et al., 2005). Positive and negative control sera were included on every plate.
8. In vitro spleen cell proliferation
Proliferation of spleen cells was performed as previously described (BONENFANT et al., 2001). Three mice of each group were sacrificed at day 62 of the experiment. Single-cell suspensions were obtained by gentle squeezing of whole spleens in erythrocyte lysis buffer (155 mM ammonium chloride, 10 mM potassium hydrogen carbonate, 0.1 mM EDTA [pH 7.4]) (Merck). Residual debris was removed by passage through a nylon gaze. The recovered cell suspension was washed once in RPMI 1640 (Life Technologies), and cells were resuspended and plated in RPMI 1640 medium (GIBCO) supplemented with 5% fetal calf serum, HEPES (25 mM; Sigma), L-glutamine (1 mM; BioWhittaker), sodium pyruvate (1 mM; Sigma), b-mercaptoethanol (50 mM), and penicillin-streptomicin (1 mM; Sigma) and seeded at 5 x 105 cells per well in triplicate in flat-bottomed 96-well microtiter plates (Costar) in 200 mL of culture medium either 1% phytohemagglutinin (PHA-M) or 5, 10, and 15 µg of rROP2 per mL. The plates were incubated in 5% CO2 at 37 ºC for 4 days. The viability of cells was higher than 85% as determined by trypan blue. Proliferating cells were determined using Vybrant MTT Cell Proliferation Assay Kit (Molecular Probes) following the manufacturer's recommendations. Absorbance measurements at 540 nm were made using a microplate reader and proliferation was expressed as total number of cells.
Results
1. Expression and purification of recombinant ROP2 protein
An 1103 bp segment (nt 1022-2125, fragment 196-561) (NIGRO et al, 2001) of the TgROP2 gene (Genbank Accession No. Z36906) was expressed as a 54 kDa polyHis fusion protein in E. coli Rosetta 2 cells (Figure 1a). Culture conditions were optimized to achieve a recombinant protein concentration equal to approx. 1 µg.mL1 culture medium. A band about 54 kDa was clearly observed in an SDS-PAGE only from soluble fraction after four hours of induction. The amount of purified ROP2 was estimated as 1 µg.mL1 (Figure 1a). Western Blots of recombinant TgROP2 protein with antibody anti-His confirmed this size estimation. (Figure 1b)
2. Immunization of mice with rROP2 induced IgG and IgA response
Anti-T. gondii IgG antibodies were seen in 5/10 (50%) mice at day 62 of the experiment (Figure 2). The mean OD in these animals was 0.307 (cut-off = 0.240). Two animals also showed IgA antibodies in serum (2/10, 20%) at day 62 and OD mean = 0.078 (cut-off = 0.101) (Figure 2).
3. Cellular proliferative response
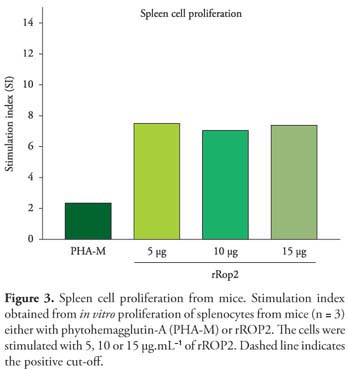
Splenocytes from mice, which received rROP2 with Quil-A, elicited a significant lymphocyte proliferation response in all three antigen concentrations. The proliferation of splenocytes revealed high stimulation indices (SI), when stimulated with 5, 10 and 15 µg of rROP2 (Figure 3). Proliferation of splenocytes higher than 6 were considered high indices of stimulation and less than 2 were considered low indices of proliferation.
Discussion
Vaccination in susceptible animals with either virulent or attenuated parasites has been a successful approach against several protozoan diseases (SCHETTERS, 1995). However, due to their potential adverse effects and the need to preserve cold-chain storage of live and attenuated vaccines, subunit vaccines derived from native antigens of the parasite or recombinant proteins from cloned DNA may overcome these difficulties (JENKINS, 2001).
Among rhoptries proteins, ROP2 has been proposed as a successful vaccine candidate (SAAVEDRA et al., 1996). It has been found that anti-ROP2 antibodies caused a decrease in invasion ability of T. gondii RH tachyzoites in vitro (MISHIMA et al., 2002). In the present study, DNA sequences encoding fragments of T. gondii ROP2 proteins were cloned and expressed under Trc promoter.
There have been some reports of expression and purification of rROP2, (JACQUET et al., 1999; NIGRO et al., 2001; IGARASHI et al., 2008a). However, in most of them rROP2 expression level has been found very low, with high degree of degradation and purification under denaturing conditions (JACQUET et al., 1999; IGARASHI et al., 2008b). Jacquet et al. (1999) constructed three different versions of rROP2 to obtain high levels of rROP2 expression. These authors found that fusion of TRX or MBP to N-terminal of ROP2 increases the expression level of this protein, however, this preparation made the protein insoluble.
The present study was similar to that of Igarashi et al. (2008a) who cloned, expressed and purified rROP2, rGRA5 and rGRA7 from T. gondii. The authors cloned a fragment of rop2 with all 1678 nucleotides into pET102/D-TOPO® vector and the recombinant protein was purified under denaturing conditions on Ni-NTA resin (Qiagen®). They found high levels of rROP2 expression in the soluble aqueous fraction and that a larger version of rTgROP2 (72 kDa) was expressed at a lower concentration (300 µg.mL1) which is considerable lower than rROP2 expression found in the present study. It is interesting that rTgROP2 in the present study had an Mr similar to native TgROP2 (54 kDa).
In the present study we focused on the expression of an rROP2 fragment which could be expressed in soluble fraction and more immunogenic than our previous study. We focused on a fragment of ROP2, which express a protein with 54 kDa and with immunogenic regions that are able to express protein from three T-cell epitopes (SAAVEDRA et al., 1996).
Cellular immune response is important for protection against T. gondii infection (JOHNSON et al., 1993). In this study, splenocytes from mice immunized with rROP2 had a strong proliferation response to these antigens when compared with positive control (phytohemagglutinin-A) that showed indices of stimulation 2. The splenocytes from mice stimulated with rROP2 showed indices higher than 6 with significant cellular immune response when stimulated with 5, 10 or 15 µg of rROP2. Protective immunity against T. gondii infection is generally cell-mediated with special involvement of CD8+ and IFN- γ (GAZZINELLI et al., 1993). INF-γ was subsequently found to activate enterocytes to render them capable of inhibiting replication of the intracellular parasite (BUZONI-GATEL et al., 1999).
In the present study only 50 and 20% of the animals showed IgG and IgA antibody levels against rROP2, respectively. This could be explained by the small number of recombinant vaccine doses (n = 3) used in the study or that there was variation in the dose delivered via intranasal route. Repeated immunizations might evoke stronger memory responses (DEBARD et al., 1996). In contrast, high levels of serum IgG antibodies were seen in mice after immunization via intranasal route with SAG1 (DEBARD et al., 1996; BONENFANT et al., 2001). Debard et al. (1996) hypothesized that the passage of SAG1 protein through the mucosa might be important for stronger serum antibody response and this could limit tachyzoite entry into the brain. There was no correlation between serum IgG levels and intestinal protection against T. gondii (FRENKEL et al., 1991). Imperfect correlations between intestinal immunity to T. gondii and circulating antibodies have also been reported (FRENKEL; DUBEY, 1972; FRENKEL et al., 1991). Additionally, Frenkel and Smith (1982) described that some cats do not develop antibodies after shedding oocysts suggesting a genetic basis to immune response.
Our choice of intranasal route of immunization is based on previous studies where intranasal injection was an effective way of providing a disseminated mucosal antibody response as well as systemic immunity (DEBARD et al., 1996). The stimulation of spleen cells after intranasal immunization revealed the existence of a common mucosal immune system. Intranasal immunization triggers both mucosal and systemic T- and B-cell responses and can be used to target pathogens that invade far from the immunization site, such as the intestines.
Natural T. gondii infection leads to strong, lifelong non-sterile protective immunity. The persistence of memory T lymphocytes, stimulated by regular rupture of T. gondii tissue cysts or by recurrent contact with infected food, prevents reinfection in most individuals (HUNTER; REICHMANN, 2001). An effective, safe vaccine should mimic the immune response seen in natural infections and protect the host against T. gondii infection. T. gondii rhoptry antigens have shown satisfactory immunogenicity seeming particularly valuable vaccine candidates for toxoplasmosis (DLUGONSKA, 2008).
An effective vaccine should protect against both acute and chronic T. gondii infection. Furthermore, mucosal immunity via intranasal route has been found to be the most relevant vaccination strategy to achieve efficient immunogenicity of nonreplicating antigens (DEBARD et al., 1996; VELGE-ROUSSEL et al., 2000).
In this study, we report the production of rROP2 in bacteria as soluble protein using the pTrcHis expression system and demonstrated that immunization via intranasal route of rROP2 plus Quil-A elicits both humoral and cellular responses in BALB/c mice.
Aknowledgment
The present article was supported by the National Council for Scientific and Technological Development (CNPq) (Grants No. 561893/2008-0, and 579843/2008-4), and Fundação Araucária (number 194/07). J.L. Garcia is recipient of CNPq fellowship.
Received May 24, 2010
Accepted November 5, 2010
- BONENFANT, C. et al. Intranasal immunization with SAG1 and nontoxic mutant heat-labile enterotoxins protects mice against Toxoplasma gondii. Infection and Immunity, v. 69, n. 3, p. 1605-1612, 2001.
- BOURGUIN, I.; CHARDÈS, T.; BOUT, D. Oral immunization with Toxoplasma gondii antigens in association with cholera toxin induces enhanced protective and cell-mediated immunity in C57BL/6 mice. Infection and Immunity, v. 61, n. 5, p. 2082-2088, 1993.
- BUZONI-GATEL, D. et al. Intraepithelial lymphocytes traffic to the intestine and enhance resistance to Toxoplasma gondii oral infection. Journal of Immunology, v. 162, n. 10, p. 5846-5852, 1999.
- COX, J. C.; COULTER, A. R. Adjuvants-A classification and review of their modes of action. Vaccine, v. 15, n. 3, p. 248-256, 1997.
- DEBARD, N.; BUZONI-GATEL, D.; BOUT, D. Intranasal immunization with SAG1 protein of Toxoplasma gondii in association with cholera toxin dramatically reduces development of cerebral cysts after oral infection. Infection and Immunity, v. 64, n. 6, p. 2158-2166, 1996.
- DLUGONSKA, H. Toxoplasma rhoptries: Unique secretory organelles and source of promising vaccine proteins for immunoprevention of toxoplasmosis. Journal of Biomedicine and Biotechnology, p. 1-7, 2008.
- FRENKEL, J. K.; DUBEY, J. P. Toxoplasmosis and its prevention in cats and man. The Jounal of Infectious Diseases, v. 126, n. 6, p. 64-673, 1972.
- FRENKEL, J. K. et al. Prospective vaccine prepared from a new mutant of Toxoplasma gondii for use in cats. American Journal of Veterinary Research, v. 52, n. 5, p. 759-763, 1991.
- FRENKEL, J. K.; SMITH, D. D. Immunization of cats against shedding of Toxoplasma oocysts. The Journal of Parasitology, v. 68, n. 5, p. 744-748, 1982.
- GARCIA, J. L. et al. Partial protection against tissue cysts formation in pigs vaccinated with crude rhoptry proteins of Toxoplasma gondii Veterinary Parasitology, v. 129, n. 3-4, p. 209-217, 2005.
- GARCIA, J. L. et al. Protective activity against oocyst shedding in cats vaccinated with crude rhoptry proteins of the Toxoplasma gondii by intranasal route. Veterinary Parasitology, v. 145, n. 3-4, p. 197-206, 2007.
- GARCIA, J. L. et al. Toxoplasma gondii: Detection by mouse bioassay, histopathology, and polymerase chain reaction in tissues from experimentally infected pigs. Experimental Parasitology, v. 113, n. 4, p. 267-271, 2006.
- GAZZINELLI, R. T.; DENKERS, E. Y.; SHER, A. Host resistance to Toxoplasma gondii: model for studying the selective induction of cell-mediated immunity by intracellular parasites. Infectious Agents and Disease, v. 2, n. 3, p. 139-149, 1993.
- HANAHAN D. Studies on transformation of Escherichia coli with plasmids. Journal Molecular Biology, v. 166, p. 557-580, 1983.
- HAUMONT, M. et al. Protective Immunity against Congenital Toxoplasmosis with recombinant SAG1 protein in a Guinea pig model. Infection and Immunity, v. 68, n. 9, p. 4948-4953, 2000.
- HIRABAYASHI, Y. et al. Comparison of intranasal inoculation of influenza HA vaccine combined with cholera toxin B subunit with oral or parenteral vaccination. Vaccine, v. 8, n. 3, p. 243-248, 1990.
- HUNTER, C. A.; REICHMANN, G. Immunology of Toxoplasma infection. In: JOYNSON, D. H.; WREGHITT, T. J. (Ed.). Toxoplasmosis A Comprehensive Clinical Guide. Cambridge University Press, 2001. p.43-57.
- IGARASHI, M. et al. Toxoplasma gondii: Evaluation of an intranasal vaccine using recombinant proteins against brain cyst formation in BALB/c mice. Experimental Parasitology, v. 118, n. 3, p. 386-392, 2008b.
- IGARASHI, M., et al. Toxoplasma gondii: clonig, sequencing, expression and antigenic characterization of ROP2, GRA5 and GRA7. Genetics and Molecular Research, v. 7, n. 2, p. 305-313, 2008a.
- JACQUET, A. et al. Expression of a recombinant Toxoplasma gondii ROP2 fragment as a fusion protein in bacteria circumvents insolubility and proteolytic degradation. Protein Expression and Purification, v. 17, n. 3, p. 392-400, 1999.
- JENKINS, M. C. Advances and prospects for subunit vaccines against protozoa of veterinary importance. Veterinary Parasitology, v. 101, n. 3-4, p. 291-310, 2001.
- JOHNSON, L. L.; VANDERVEGT, F. P.; HAVELL, E. A. Gamma interferon-dependent temporary resistance to acute Toxoplasma gondii infection independent of CD4+ or CD8+ lymphocytes. Infection and Immunity, v. 61, n. 12, p. 5174-5180, 1993.
- MARTIN, V. et al. Recombinant GRA4 or ROP2 Protein Combined with Alum or the gra4 Gene Provides Partial Protection in Chronic Murine Models of Toxoplasmosis Clinical And Diagnostic Laboratory Immunology, v. 11, n. 4, p. 704-710, 2004.
- MISHIMA, M. et al. Recombinant feline herpesvirus type 1 expressing Toxoplasma gondii ROP2 antigen inducible protective immunity in cats. Parasitology Research, v. 88, n. 2, p. 144-149, 2002.
- NIGRO, M. et al. High level of expression of the Toxoplasma gondii-recombinant Rop2 protein in Escherichia coli as a soluble form for optimal use in diagnosis. Molecular Biotechnology, v. 18, n. 3, p. 269-73, 2001.
- ROBERTS, C. W.; ALEXANDER, J. Studies on a murine model of congenital toxoplasmosis: vertical disease transmission only occurs in BALB/c mice infected for the first time during pregnancy. Parasitology, v. 104, n. 1, p. 19-23, 1992.
- SAAVEDRA, R. et al. Epitopes recognized by human T lymphocytes in the ROP2 protein antigen of Toxoplasma gondii. Infection and Immunity, v. 64, n. 9, p. 3858-3862, 1996.
- SCHETTERS, T. Vaccine development from a commercial point of view. Veterinary Parasitology, v. 57, p. 267-275, 1995.
- SMITH, R. E.; DONACHIE, A. M.; MOWAT, A. M. Immune stimulating complexes as mucosal vaccines. Immunology and Cell Biology, v. 76, p. 263-269, 1998.
- VELGE-ROUSSEL, F. et al. Intranasal immunization with Toxoplasma gondii SAG1 induces protective cells into both NALT and GALT compartments. Infection and Immunity, v. 68, n. 2, p. 969-972, 2000.
- WU, H. Y.; RUSSEL, M. W. Nasal lymphoid tissue, intranasal immunization, and compartmentalization of the common mucosal immune system. Immunologic Research, v. 16, n. 2, p. 187-201, 1997.
Corresponding author:
Publication Dates
-
Publication in this collection
07 Feb 2011 -
Date of issue
Dec 2010
History
-
Received
24 May 2010 -
Accepted
05 Nov 2010