Resumos
Abstract
Molecular phylogenetic analyses based on ETS, ITS and atpB - rbcL spacer sequences assessed the phylogenetic status of the southern Brazil sea asparagus species of the genus Salicornia (Salicornioideae, Amaranthaceae). Accessions of Patos Lagoon estuary (32° S) were obtained from wild plants and two pure line lineages, selected from contrasting prostrate (BTH1) and decumbent (BTH2) ecomorphotypes found locally. Patos Lagoon wild plants, BTH1 and BTH2 f4 progenies showed 100% identical sequences for the atpB - rbcL and ITS spacers, only two mutations for ETS. Comparison of the sequences of these three markers with GenBank records confirmed the identity of Brazilian accessions as Salicornia neei . Maximum-likelihood phylogenetic analysis of ETS sequences indicated that the southern Brazilian accessions of Salicornia certainly are not close to any of the Salicornia ambigua accessions in GenBank, which are restricted to the northern hemisphere, nor are they related to any Salicornia fruticosa/Salicornia perennis clade accessions, which are also restricted to Eurasia. All above cited species have been wrongly applied to the southern Brazil sea asparagus.
Palavras-chave:
sequências de DNA; Salicornioideae; Sarcocornia; halófitas; marismas
Resumo
Análises filogenéticas moleculares baseadas em sequências das regiões espaçadoras ETS, ITS and atpB - rbcL avaliaram o status filogenético da espécie de aspargo marinho do gênero Salicornia (Salicornioideae, Amaranthaceae) presente no sul do Brasil. Os acessos do estuário da Lagoa dos Patos (32° S) foram obtidos de uma população selvagem e de duas linhagens puras de aspargo marinho, selecionadas de ecomorfotipos contrastantes com forma de crescimento prostrada (BTH1) e decumbente (BTH2) encontrados localmente. Plantas selvagens do estuário da Lagoa dos Patos e as progênies f4 de BTH1 e BTH2 mostraram sequencias 100% idênticas para a região espaçadora atpB - rbcL e ITS, apenas duas mutações em ETS. A comparação das sequências desses três marcadores com registros do GenBank confirmou a identidade dos acessos como Salicornia neei . Análise filogenética pelo método de máxima verossimilhança das sequências ETS indicou que os acessos de Salicornia do sul do Brasil certamente não são semelhantes a nenhum acesso de Salicornia ambigua no GenBank, os quais possuem distribuição restrita ao hemisfério norte, e nem são relacionados com qualquer acesso do clado Salicornia fruticosa/Salicornia perennis, este último restrito a Eurásia. Todas as espécies citadas acima têm sido erroneamente consideradas como sinonímias para o aspargo marinho do sul do Brasil.
Palavras-chave:
sequências de DNA; Salicornioideae; Sarcocornia; halófitas; marismas
Introduction
The small succulent shrubs with leafless stems and branches of the genera Salicornia L. and Sarcocornia A.J.Scott (Salicornioideae, Amaranthaceae) are characterized as ‟extreme halophytes” based on their ability to thrive in seawater-flooded and hypersaline soils (Davy et al. 2006Davy AJ, Bishop GF, Mossman H, Redondo-Goméz S, Castillo JM, Castellanos EM, Luque T & Figueroa ME (2006) Sarcocornia perennis (Miller) A.J.Scott. Biological Flora of the British Isles, n. 244. Journal of Ecology 94: 1035-1048. DOI: 10.1111/j.1365-2745.2006.01156.x
https://doi.org/10.1111/j.1365-2745.2006...
; Steffen et al. 2015Steffen S, Ball P, Mucina L & Kadereit G (2015) Phylogeny, biogeography and ecological diversification of Sarcocornia (Salicornioideae, Amaranthaceae). Annals of Botany 115: 353-368. DOI: 10.1093/aob/mcu260
https://doi.org/10.1093/aob/mcu260...
). These halophytes have been consumed by humans for centuries, and to date are cultivated with a broad range of saline water sources and sold as ‘sea asparagus’ or ‘samphire’, particularly used as functional gourmet food, animal feed meal and oils for biodiesel (Ventura & Sagi 2013Ventura Y & Sagi M (2013) Halophyte crop cultivation: the case of Salicornia and Sarcocornia. Environmental and Experimental Botany 92:144-153.; Ventura et al. 2015Ventura Y, Eshel A & Pasternak D & Sagi M (2015) The development of halophyte-based agriculture: past and present. Annals of Botany 115:327-331.; Costa & Herrera 2016Costa CSB & Herrera OB (2016) Halophytic life in Brazilian salt flats: biodiversity, uses and threats. In: Khan MA, Boër B, Azturk M, Clüsener-Godt M, Gul B & Breckle S-W (eds.) Sabkha ecosystems Vol. V. The Americas. Springer, Berlin. Pp. 11-27. DOI: 10.1007/978-3-319-27093-7_2
https://doi.org/10.1007/978-3-319-27093-...
).
The paucity of characters due to the strongly reduced morphology of Salicornia and Sarcocornia, the great phenotypic plasticity (Kadereit et al. 2006Kadereit G, Mucina L & Freitag H (2006) Phylogeny of Salicornioideae (Chenopodiaceae): diversification, biogeography, and evolutionary trends in leaf and flower morphology. Taxon 55: 617-642. DOI: 10.2307/25065639
https://doi.org/10.2307/25065639...
; Alonso & Crespo 2008Alonso MA & Crespo MB (2008) Taxonomic and nomenclatural notes on South American taxa of Sarcocornia A.J. Scott (Chenopodiaceae). Annales Botanici Fennici 45: 241-254. DOI: 10.5735/085.045.0401
https://doi.org/10.5735/085.045.0401...
; Doncato & Costa 2018Doncato KB & Costa CSB (2018) Growth and mineral composition of two lineages of the sea asparagus Sarcocornia ambigua irrigated with shrimp farm saline effluent. Experimental Agriculture 54: 399-416. DOI: 10.1017/S0014479717000096
https://doi.org/10.1017/S001447971700009...
) and high intrapopulation genetic diversity in addition to the formation of hybrids between sympatric species through polyploidy (Davy et al. 2006Davy AJ, Bishop GF, Mossman H, Redondo-Goméz S, Castillo JM, Castellanos EM, Luque T & Figueroa ME (2006) Sarcocornia perennis (Miller) A.J.Scott. Biological Flora of the British Isles, n. 244. Journal of Ecology 94: 1035-1048. DOI: 10.1111/j.1365-2745.2006.01156.x
https://doi.org/10.1111/j.1365-2745.2006...
; Steffen et al. 2015Steffen S, Ball P, Mucina L & Kadereit G (2015) Phylogeny, biogeography and ecological diversification of Sarcocornia (Salicornioideae, Amaranthaceae). Annals of Botany 115: 353-368. DOI: 10.1093/aob/mcu260
https://doi.org/10.1093/aob/mcu260...
) generate a notorious difficult taxonomy making it almost impossible for non-specialists to determine most species of these genera. Molecular makers have been used to clarify the phylogeny and biogeography of the subfamily Salicornioideae (Kadereit et al. 2006Kadereit G, Mucina L & Freitag H (2006) Phylogeny of Salicornioideae (Chenopodiaceae): diversification, biogeography, and evolutionary trends in leaf and flower morphology. Taxon 55: 617-642. DOI: 10.2307/25065639
https://doi.org/10.2307/25065639...
; Steffen et al. 2015Steffen S, Ball P, Mucina L & Kadereit G (2015) Phylogeny, biogeography and ecological diversification of Sarcocornia (Salicornioideae, Amaranthaceae). Annals of Botany 115: 353-368. DOI: 10.1093/aob/mcu260
https://doi.org/10.1093/aob/mcu260...
; Piirainen et al. 2017Piirainen M, Liebisch O & Kadereit G (2017) Phylogeny, biogeography, systematics and taxonomy of Salicornioideae (Amaranthaceae/Chenopodiaceae) - A cosmopolitan, highly specialized hygrohalophyte lineage dating back to the Oligocene. Taxon 66: 109-132. DOI: 10.12705/661.6
https://doi.org/10.12705/661.6...
). In a molecular phylogeny based on Internal Transcribed Spacer (ITS) and atp B- rbc L spacer (chloroplast DNA) sequences of Sarcocornia formed a monophyletic lineage together with the genus Salicornia, which is clearly separated from the genus Arthrocnemum (R.Br.) Moq. (Kadereit et al. 2006Kadereit G, Mucina L & Freitag H (2006) Phylogeny of Salicornioideae (Chenopodiaceae): diversification, biogeography, and evolutionary trends in leaf and flower morphology. Taxon 55: 617-642. DOI: 10.2307/25065639
https://doi.org/10.2307/25065639...
). External Transcribed Spacer (ETS) and matK-trnK intron (chloroplast DNA) sequence data showed that Salicornia is clearly nested in Sarcocornia and sister to a clade comprising American and Eurasian Sarcocornia (Kadereit et al. 2007Kadereit G, Ball P, Beer S, Mucina L, Sokoloff D, Teege P, Yaprak AE & Freitag H (2007.) A taxonomic nightmare comes true: phylogeny and biogeography of glassworts (Salicornia L., Chenopodiaceae). Taxon 56: 1143-1170. DOI: 10.2307/25065909
https://doi.org/10.2307/25065909...
; Piirainen et al. 2017Piirainen M, Liebisch O & Kadereit G (2017) Phylogeny, biogeography, systematics and taxonomy of Salicornioideae (Amaranthaceae/Chenopodiaceae) - A cosmopolitan, highly specialized hygrohalophyte lineage dating back to the Oligocene. Taxon 66: 109-132. DOI: 10.12705/661.6
https://doi.org/10.12705/661.6...
). In the latest worldwide molecular phylogenetic treatment of Salicornioideae, Piirainen et al. (2017)Piirainen M, Liebisch O & Kadereit G (2017) Phylogeny, biogeography, systematics and taxonomy of Salicornioideae (Amaranthaceae/Chenopodiaceae) - A cosmopolitan, highly specialized hygrohalophyte lineage dating back to the Oligocene. Taxon 66: 109-132. DOI: 10.12705/661.6
https://doi.org/10.12705/661.6...
showed that Sarcocornia is paraphyletic in relation to Salicornia and proposed a new taxonomic classification, merging of Sarcocornia in three subgeneraunder Salicornia affecting 19 new nomenclatural combinations and one replacement name. In the present study, we followed the nomenclature proposed in Piirainen et al. (2017).
In the most recent taxonomic classification of the genus Salicornia in South America based on morphological features, Alonso & Crespo (2008) accepted five perennial species of sea asparagus (originally cited as Sarcocornia) and cited the synonyms and types for the accepted taxa: Salicornia ambigua Michx., S. andina Phil., S. magellanica Phil., S. neei Lag. and S. pulvinata R.E.Fr. Alonso et al. (2017)Alonso MA, Crespo MB & Freitag H (2017) Salicornia cuscoensis (Amaranthaceae/Chenopodiaceae), a new species from Peru (South America). Phytotaxa 319: 254-262. DOI: 10.11646/phytotaxa.319.3.4
https://doi.org/10.11646/phytotaxa.319.3...
described a sixth perennial species named Salicornia cuscoensis Gutte & G.K. Müll. found in high Andean saltmarshes in Peru, the identity of which is supported by differences in morphological characters and molecular analyses. Previously, Scott (1977)Scott AJ (1977) Reinstatement and revision of Salicorniaceae J.Agardh (Caryophyllales). Botanical Journal of the Linnean Society 75: 255-307. DOI: 10.1111/j.1095-8339.1977.tb01493.x
https://doi.org/10.1111/j.1095-8339.1977...
recognized only two species of perennial Salicornia in South America ( S. fruticosa and S. pulvinata) . Similar annual Salicornia plants, these two species have leafless succulent stems but distinctly unequal sized flowers in their cymes with a triangular arrangement.
Salicornia ambigua is the only species of sea asparagus listed for Brazil (Alonso & Crespo 2008Alonso MA, Crespo MB & Freitag H (2017) Salicornia cuscoensis (Amaranthaceae/Chenopodiaceae), a new species from Peru (South America). Phytotaxa 319: 254-262. DOI: 10.11646/phytotaxa.319.3.4
https://doi.org/10.11646/phytotaxa.319.3...
). It was regarded as a synonym for Salicornia gaudichaudiana Moq., Sarcocornia ambigua (Michx.) M.A.Alonso & M.B.Crespo, the Mediterranean species Sarcocornia fruticosa (L.) A.J.Scott (Scott 1977)Scott AJ (1977) Reinstatement and revision of Salicorniaceae J.Agardh (Caryophyllales). Botanical Journal of the Linnean Society 75: 255-307. DOI: 10.1111/j.1095-8339.1977.tb01493.x
https://doi.org/10.1111/j.1095-8339.1977...
, Salicornia fruticosa L. (BFG 2018) and Salicornia perennis Mill. (= Sarcocornia perennis (Miller) A.J.Scott) (Costa et al. 2006Costa CSB, Armstrong R, Detrés Y, Koch EW, Bertiller M, Beeskow A, Neves LS, Tourn GM, Bianciotto OA, Pinedo LB, Blessio AY & San Roman N (2006)Effect of ultraviolet-B radiation on salt marsh vegetation: trends of the genus Salicornia along the Americas. Photochemical and Photobiology 82: 878-886. DOI: 10.1562/2005-10-30-RA-729
https://doi.org/10.1562/2005-10-30-RA-72...
; Isacch et al. 2006Isacch JP, Costa CSB, Rodríguez-Gallegol L, Conde D, Escapa M, Gagliardini DA & Iribarne O (2006)Distribution of salt marshes plant communities associated with environmental factors along a latitudinal gradient on the south-east Atlantic coast. Journal of Biogeography 33: 888-900. DOI: 10.1111/j.1365-2699.2006.01461.x
https://doi.org/10.1111/j.1365-2699.2006...
; Davy et al. 2006Davy AJ, Bishop GF, Mossman H, Redondo-Goméz S, Castillo JM, Castellanos EM, Luque T & Figueroa ME (2006) Sarcocornia perennis (Miller) A.J.Scott. Biological Flora of the British Isles, n. 244. Journal of Ecology 94: 1035-1048. DOI: 10.1111/j.1365-2745.2006.01156.x
https://doi.org/10.1111/j.1365-2745.2006...
) or the North American Salicornia virginica L. (Reitz 1961Reitz PR (1961) Vegetação da zona marítima de Santa Catarina. Sellowia 13: 17-115.). Additionally, many plants referred to Salicornia ambigua Michx. and Salicornia fruticosa (= Sarcocornia fruticosa) on the coasts of Uruguay and NE Argentina are indistinguishable from southern Brazilian sea asparagus (Costa & Davy 1992Costa CSB & Davy AJ (1992) Coastal salt marsh communities of Latin America. In: Seeliger U (ed.) Coastal plant communities of Latin America. Academic Press, New York. Pp. 179-199. DOI: 10.1016/B978-0-08-092567-7.50018-0
https://doi.org/10.1016/B978-0-08-092567...
; Davy et al. 2006Davy AJ, Bishop GF, Mossman H, Redondo-Goméz S, Castillo JM, Castellanos EM, Luque T & Figueroa ME (2006) Sarcocornia perennis (Miller) A.J.Scott. Biological Flora of the British Isles, n. 244. Journal of Ecology 94: 1035-1048. DOI: 10.1111/j.1365-2745.2006.01156.x
https://doi.org/10.1111/j.1365-2745.2006...
; Isacch et al. 2006Isacch JP, Costa CSB, Rodríguez-Gallegol L, Conde D, Escapa M, Gagliardini DA & Iribarne O (2006)Distribution of salt marshes plant communities associated with environmental factors along a latitudinal gradient on the south-east Atlantic coast. Journal of Biogeography 33: 888-900. DOI: 10.1111/j.1365-2699.2006.01461.x
https://doi.org/10.1111/j.1365-2699.2006...
). Steffen et al. (2015)Steffen S, Ball P, Mucina L & Kadereit G (2015) Phylogeny, biogeography and ecological diversification of Sarcocornia (Salicornioideae, Amaranthaceae). Annals of Botany 115: 353-368. DOI: 10.1093/aob/mcu260
https://doi.org/10.1093/aob/mcu260...
using molecular phylogenetic analyses based on ETS, atp B- rbc L and rpl 32- trn L characterized S. fruticosa and S. perennis as exclusively Eurasian and partially supported Alonso & Crespo (2008)Alonso MA & Crespo MB (2008) Taxonomic and nomenclatural notes on South American taxa of Sarcocornia A.J. Scott (Chenopodiaceae). Annales Botanici Fennici 45: 241-254. DOI: 10.5735/085.045.0401
https://doi.org/10.5735/085.045.0401...
classification of South American Salicornia, but no accessions from the Brazilian coast was assessed.
Although both morphological (Scott 1977Scott AJ (1977) Reinstatement and revision of Salicorniaceae J.Agardh (Caryophyllales). Botanical Journal of the Linnean Society 75: 255-307. DOI: 10.1111/j.1095-8339.1977.tb01493.x
https://doi.org/10.1111/j.1095-8339.1977...
; Alonso & Crespo 2008Alonso MA & Crespo MB (2008) Taxonomic and nomenclatural notes on South American taxa of Sarcocornia A.J. Scott (Chenopodiaceae). Annales Botanici Fennici 45: 241-254. DOI: 10.5735/085.045.0401
https://doi.org/10.5735/085.045.0401...
) and molecular studies (Steffen et al. 2015Steffen S, Ball P, Mucina L & Kadereit G (2015) Phylogeny, biogeography and ecological diversification of Sarcocornia (Salicornioideae, Amaranthaceae). Annals of Botany 115: 353-368. DOI: 10.1093/aob/mcu260
https://doi.org/10.1093/aob/mcu260...
; Piirainen et al. 2017Piirainen M, Liebisch O & Kadereit G (2017) Phylogeny, biogeography, systematics and taxonomy of Salicornioideae (Amaranthaceae/Chenopodiaceae) - A cosmopolitan, highly specialized hygrohalophyte lineage dating back to the Oligocene. Taxon 66: 109-132. DOI: 10.12705/661.6
https://doi.org/10.12705/661.6...
) recognized the occurrence of Salicornia ambigua in Americas, they worked with specimens from distinct locations and poorly sampled the Brazilian coast. Alonso & Crespo (2008)Alonso MA & Crespo MB (2008) Taxonomic and nomenclatural notes on South American taxa of Sarcocornia A.J. Scott (Chenopodiaceae). Annales Botanici Fennici 45: 241-254. DOI: 10.5735/085.045.0401
https://doi.org/10.5735/085.045.0401...
recognized Brazilian plants as S. ambigua based on an over 45 years-old herbarium specimen with slender woody stems at the base, and long and narrow terminal inflorescence that produced small seeds with short appressed hairs mostly on the edge. Their sampling of S. ambigua pointed out the southernmost records to Mar del Plata in the Argentinian province of Buenos Aires (39° S), and they also examined specimens of this taxon collected in the Caribbean region of Dominican Republic (19° N). The original description of “ Salicornia ambigua ” was from plants from North American “Carolinas” (Michaux, 1803 sec. Alonso & Crespo 2008Alonso MA & Crespo MB (2008) Taxonomic and nomenclatural notes on South American taxa of Sarcocornia A.J. Scott (Chenopodiaceae). Annales Botanici Fennici 45: 241-254. DOI: 10.5735/085.045.0401
https://doi.org/10.5735/085.045.0401...
) and Steffen et al. (2015)Steffen S, Ball P, Mucina L & Kadereit G (2015) Phylogeny, biogeography and ecological diversification of Sarcocornia (Salicornioideae, Amaranthaceae). Annals of Botany 115: 353-368. DOI: 10.1093/aob/mcu260
https://doi.org/10.1093/aob/mcu260...
highlighted that in Florida S. ambigua occurs in extensive tidal flats. The S. ambigua accessions analyzed in Steffen et al. (2015) were collected from the Caribbean region and the east Atlantic coast of U.S.A, and plants from Atlantic coast of Argentina turned out to belong to the S. neei - S. pacifica - S. magellanica lineage. Alonso & Crespo (2008)Alonso MA & Crespo MB (2008) Taxonomic and nomenclatural notes on South American taxa of Sarcocornia A.J. Scott (Chenopodiaceae). Annales Botanici Fennici 45: 241-254. DOI: 10.5735/085.045.0401
https://doi.org/10.5735/085.045.0401...
also sampled and listed S. neei and S. magellanica for the Atlantic coast of South America. Salicornia magellanica that would be endemic to southern Argentina and southern Chile, and the widely spread S. neei found southwards of Buenos Aires province and saline continental lowlands of Argentina (except for Strait of Magellan area), as well as in the Pacific coast of Chile and Peru. In their molecular study Steffen et al. (2015) recognized the phylogenetic affinity of these two species and their occurrence in the area described by Alonso & Crespo (2008)Alonso MA & Crespo MB (2008) Taxonomic and nomenclatural notes on South American taxa of Sarcocornia A.J. Scott (Chenopodiaceae). Annales Botanici Fennici 45: 241-254. DOI: 10.5735/085.045.0401
https://doi.org/10.5735/085.045.0401...
.
The taxonomy and the geographic distribution of perennial Salicornia species in the Atlantic coast of South America are still controversial. The morphological identification of dried herbarium specimens can be very difficult, and the Brazilian coast represents a biogeographic gap in molecular studies of Salicornioideae in Americas. The Brazilian sea asparagus of the genus Salicornia shows a recent history of successful cultivation with saline water and shrimp farm effluent (Costa & Herrera 2016Costa CSB & Herrera OB (2016) Halophytic life in Brazilian salt flats: biodiversity, uses and threats. In: Khan MA, Boër B, Azturk M, Clüsener-Godt M, Gul B & Breckle S-W (eds.) Sabkha ecosystems Vol. V. The Americas. Springer, Berlin. Pp. 11-27. DOI: 10.1007/978-3-319-27093-7_2
https://doi.org/10.1007/978-3-319-27093-...
), high nutritional quality (Bertin et al. 2014Bertin RL, Gonzag LV, Borges GSC, Azevedo MS, Maltez HF, Heller M, Micke GA, Tavares LBB & Fett R (2014) Nutrient composition and, identification/quantification of major phenolic compounds in Sarcocornia ambigua (Amaranthaceae) using HPLC-ESI-MS/MS. Food Research International 55: 404-411. DOI:10.1016/j.foodres.2013.11.036
https://doi.org/0.1016/j.foodres.2013.11...
; Costa et al. 2014)Costa CSB, Vicenti JRM, Morón-Villarreyes JA, Caldas S, Cardoso LV, Freitas RF & D’oca MGM (2014) Extraction and characterization of lipids from Sarcocornia ambigua meal: a halophyte biomass produced with shrimp farm effluent irrigation. Anais da Academia Brasileira de Ciências 86: 935-943. DOI: 10.1590/0001-3765201420130022
https://doi.org/10.1590/0001-37652014201...
and chemical characteristics for biofuel production (D’Oca et al. 2012D’oca MGM, Moron-Villarreyes JA, Lemoes JS & Costa CSB (2012) Fatty acids composition in seeds of the South American glasswort Sarcocornia ambigua. Anais da Academia Brasileira de Ciências 84: 865-870. DOI: 10.1590/S0001-37652012005000044
https://doi.org/10.1590/S0001-3765201200...
; Costa et al. 2014Costa CSB, Vicenti JRM, Morón-Villarreyes JA, Caldas S, Cardoso LV, Freitas RF & D’oca MGM (2014) Extraction and characterization of lipids from Sarcocornia ambigua meal: a halophyte biomass produced with shrimp farm effluent irrigation. Anais da Academia Brasileira de Ciências 86: 935-943. DOI: 10.1590/0001-3765201420130022
https://doi.org/10.1590/0001-37652014201...
). However the designation of plants found in Brazil as Salicornia ambigua (= Sarcocornia ambigua) is uncertain. The proper taxonomic identification and the establishment genetically defined lines of the Brazilian sea asparagus of the genus Salicornia are necessary for the development of breeding programs that produce plants with desirable agricultural traits. Using molecular phylogenetic analyses based on ETS, ITS and atpB - rbcL spacer sequences, the present study is aimed at clarifying the phylogenetic status of the southern Brazil Salicornia population by comparison with GenBank records.
Materials and Methods
Plant material
The wild plant accessions of Salicornia were collected in the salt marsh of Pólvora Island (32º01’ S, 52º06’ W) at Patos Lagoon estuary (located in the state of Rio Grande do Sul, of the southern Brazil) and 8 weeks-old accessions of f4 progenies of the Salicornia lineages BTH1 and BTH2 were obtained from the germplasm of Laboratório de Biotecnologia de Halófitas (Instituto de Oceanografia, FURG, Rio Grande, RS, Brazil). These two lineages represent contrasting prostrate and decumbent plants with reddish and green colorations, respectively, obtained by a 5 year breeding program based on pure line selection from ecomorphotypes found in local estuarine habitats subject to distinctive flooding regimes and salt stresses (Freitas & Costa 2014Freitas RF & Costa CSB (2014) Germination responses to salt stress of two intertidal populations of the perennial glasswort Sarcocornia ambigua. Aquatic Botany 117: 12-17. DOI: 10.1016/j.aquabot.2014.04.002
https://doi.org/10.1016/j.aquabot.2014.0...
; Doncato & Costa 2018Doncato KB & Costa CSB (2018) Growth and mineral composition of two lineages of the sea asparagus Sarcocornia ambigua irrigated with shrimp farm saline effluent. Experimental Agriculture 54: 399-416. DOI: 10.1017/S0014479717000096
https://doi.org/10.1017/S001447971700009...
). Since decumbent-prostrate growth forms found within different lineages of other Salicornia species might represent different stages of speciation (Steffen et al. 2015Steffen S, Ball P, Mucina L & Kadereit G (2015) Phylogeny, biogeography and ecological diversification of Sarcocornia (Salicornioideae, Amaranthaceae). Annals of Botany 115: 353-368. DOI: 10.1093/aob/mcu260
https://doi.org/10.1093/aob/mcu260...
), the DNA analysis of BTH1 and BTH2 lineages was carried out in order to assess the range of genetic variation found in southern Brazil Salicornia populations. Vegetative stem segments harvested from the three accession groups (wild, BTH1 and BTH2) were separated in situ, placed in vials, frozen in liquid nitrogen, and kept at -80 ºC until DNA extraction.
DNA extraction and DNA sequences
Total DNA was extracted from 50 mg of frozen stem material using the DNeasy Plant Mini Kit (Qiagen, Valencia, CA, USA) following a conventional hexadecyl-trimethyl-ammonium bromide (CTAB) DNA extraction protocol (Doyle 1991Doyle JJ (1991) DNA protocols for plants. In: Hewitt G, Johnson AWB & Young JPW (eds.) Molecular techniques in taxonomy. NATO ASI Series H, Cell Biology. Vol. 57. Springer-Verlag, Berlin. Pp. 283-293. DOI: 0.1007/978-3-642-83962-7_18
https://doi.org/0.1007/978-3-642-83962-7...
). PCR was carried out in a T-Professional or T-Gradient Thermocycler (Biometra, Göttingen, Germany). We used the same primer sequences, PCR recipe and cycler programme for each marker applied by Steffen et al. (2015)Steffen S, Ball P, Mucina L & Kadereit G (2015) Phylogeny, biogeography and ecological diversification of Sarcocornia (Salicornioideae, Amaranthaceae). Annals of Botany 115: 353-368. DOI: 10.1093/aob/mcu260
https://doi.org/10.1093/aob/mcu260...
. We selected one variable chloroplast (cp) marker, atpB - rbcL spacer, and ITS and ETS ribosomal DNA markers as the most variable nuclear markers known for Salicornioideae (Kadereit et al. 2007Kadereit G, Ball P, Beer S, Mucina L, Sokoloff D, Teege P, Yaprak AE & Freitag H (2007.) A taxonomic nightmare comes true: phylogeny and biogeography of glassworts (Salicornia L., Chenopodiaceae). Taxon 56: 1143-1170. DOI: 10.2307/25065909
https://doi.org/10.2307/25065909...
, 2012;
Steffen et al. 2015Steffen S, Ball P, Mucina L & Kadereit G (2015) Phylogeny, biogeography and ecological diversification of Sarcocornia (Salicornioideae, Amaranthaceae). Annals of Botany 115: 353-368. DOI: 10.1093/aob/mcu260
https://doi.org/10.1093/aob/mcu260...
;
Piirainen et al. 2017Piirainen M, Liebisch O & Kadereit G (2017) Phylogeny, biogeography, systematics and taxonomy of Salicornioideae (Amaranthaceae/Chenopodiaceae) - A cosmopolitan, highly specialized hygrohalophyte lineage dating back to the Oligocene. Taxon 66: 109-132. DOI: 10.12705/661.6
https://doi.org/10.12705/661.6...
). For details of primer sequences see Kadereit et al. (2006)Kadereit G, Mucina L & Freitag H (2006) Phylogeny of Salicornioideae (Chenopodiaceae): diversification, biogeography, and evolutionary trends in leaf and flower morphology. Taxon 55: 617-642. DOI: 10.2307/25065639
https://doi.org/10.2307/25065639...
and Steffen et al. (2015)Steffen S, Ball P, Mucina L & Kadereit G (2015) Phylogeny, biogeography and ecological diversification of Sarcocornia (Salicornioideae, Amaranthaceae). Annals of Botany 115: 353-368. DOI: 10.1093/aob/mcu260
https://doi.org/10.1093/aob/mcu260...
. PCR products were checked on 1 % agarose gels and purified subsequently using the NucleoSpin Gel and PCR clean-up-Kit (Macherey-Nagel, Düren, Germany) following the manufacturer’s manual. DNA sequences were obtained using the Big Dye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, Foster City, CA, USA) in combination with the primers mentioned above following a purification step using an Illustra SephadexTM G-50 Fine DNA Grade (GE Healthcare, Little Chalfont, UK). DNA fragments were sequenced using an automatic capillary sequencer GA3130XL (Applied Biosystems) following the Sanger method. Forward and reverse sequences were edited and merged to consensus sequences, which then were aligned using Sequencher 4.1.4 (Gene Codes Corp., Ann Arbor, MI, USA). All alignments were checked and corrected manually. ITS, atpB-rbcl spacer and ETS sequences of Brazilian accessions have been deposited in the GenBank database (accession numbers; BTH1= MG579955-MG579957;
BTH2= MG579958-MG579960;
Wild= MG579961- MG579963).
Data analysis
The chloroplast data set consisting of the atpB - rbcL spacer and the nuclear data set consisting of ITS (ITS-28S forward and ITS-18S reverse) and ETS sequences were analysed separately. Since the cp and ITS data were mostly uninformative among Salicornia samples from southern Brazil, phylogenetic analysis was performed using only ETS data. Maximum-likelihood (ML) phylogenetic analysis including bootstrapping was performed using MEGA7 (Kumar et al. 2016Kumar S, Stecher G & Tamura K (2016) MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Molecular Biology and Evolution 33: 1870-1874. DOI: 10.1093/molbev/msw074
https://doi.org/10.1093/molbev/msw074...
). The ETS analysis involved 46 nucleotide sequences. There were a total of 503 positions in the final dataset. Bootstrap support was calculated using ten random additions of taxa in 1000 bootstrap replicates and bootstrap reliability percentages of the clusters descending from each node of the tree were also calculated. We found T92 (substitution model parameters Tamura 3) with a Bayesian Information Criterion (BIC) score equal to 21142.9, transition/transversion bias (R) equal to 0.66, and nucleotide substitution rates ranging between 0.072 and 0.104 as the best evolutive model to our sequences.
Besides the 3 accessions from southern Brazil, we included DNA sequences of all coastal species of perennial Salicornia found in Americas, S. perennis and S. fruticosa (previously cited as synonyms for S. ambigua) , South African Salicornia species (S. natalensis, S. pillansii, S. tegetaria and S. capensis) and, representative species of annual Salicornia, obtained from different studies through their GenBank records and available from Steffen et al. (2015)Steffen S, Ball P, Mucina L & Kadereit G (2015) Phylogeny, biogeography and ecological diversification of Sarcocornia (Salicornioideae, Amaranthaceae). Annals of Botany 115: 353-368. DOI: 10.1093/aob/mcu260
https://doi.org/10.1093/aob/mcu260...
. Representatives of Microcnemum Ung.-Sternb., Arthrocaulon Piirainen & G. Kadereit (~ Arthrocnemum Moq.) and Tecticornia, were included as outgroups (Kadereit et al. 2006Kadereit G, Mucina L & Freitag H (2006) Phylogeny of Salicornioideae (Chenopodiaceae): diversification, biogeography, and evolutionary trends in leaf and flower morphology. Taxon 55: 617-642. DOI: 10.2307/25065639
https://doi.org/10.2307/25065639...
; Steffen et al. 2015). Altogether 46 accessions were included in the phylogenetic analysis representing approx. 10 species of perennial Salicornia, approx. 5 species of annual Salicornia and 4 outgroups (Tab. 1).
List of sequenced accessions of external transcribed spacer (ETS) with lab code, voucher information, life form (p= perennial; a= annual) and GenBank accession numbers.
Results
Accessions of Patos Lagoon wild plant, BTH1 and BTH2 f4 progenies showed 100% identical sequences for atpB - rbcL and ITS spacers, which included 709 and 647 aligned nucleotide positions, respectively (Tab. 2). The atpB - rbcL spacer and ITS sequences are nearly identical to published sequences of S. neei/S. magellanica clade, with the highest score for S. neei.
Number of aligned nucleotides, aligned position of detected mutations and mutated nucleotide sequences for three markers of southern Brazilian accessions of Salicornia (wild of the Pólvora Island, BTH1 and BTH2). Positions of variable nucleotides among accessions in italic and bold.
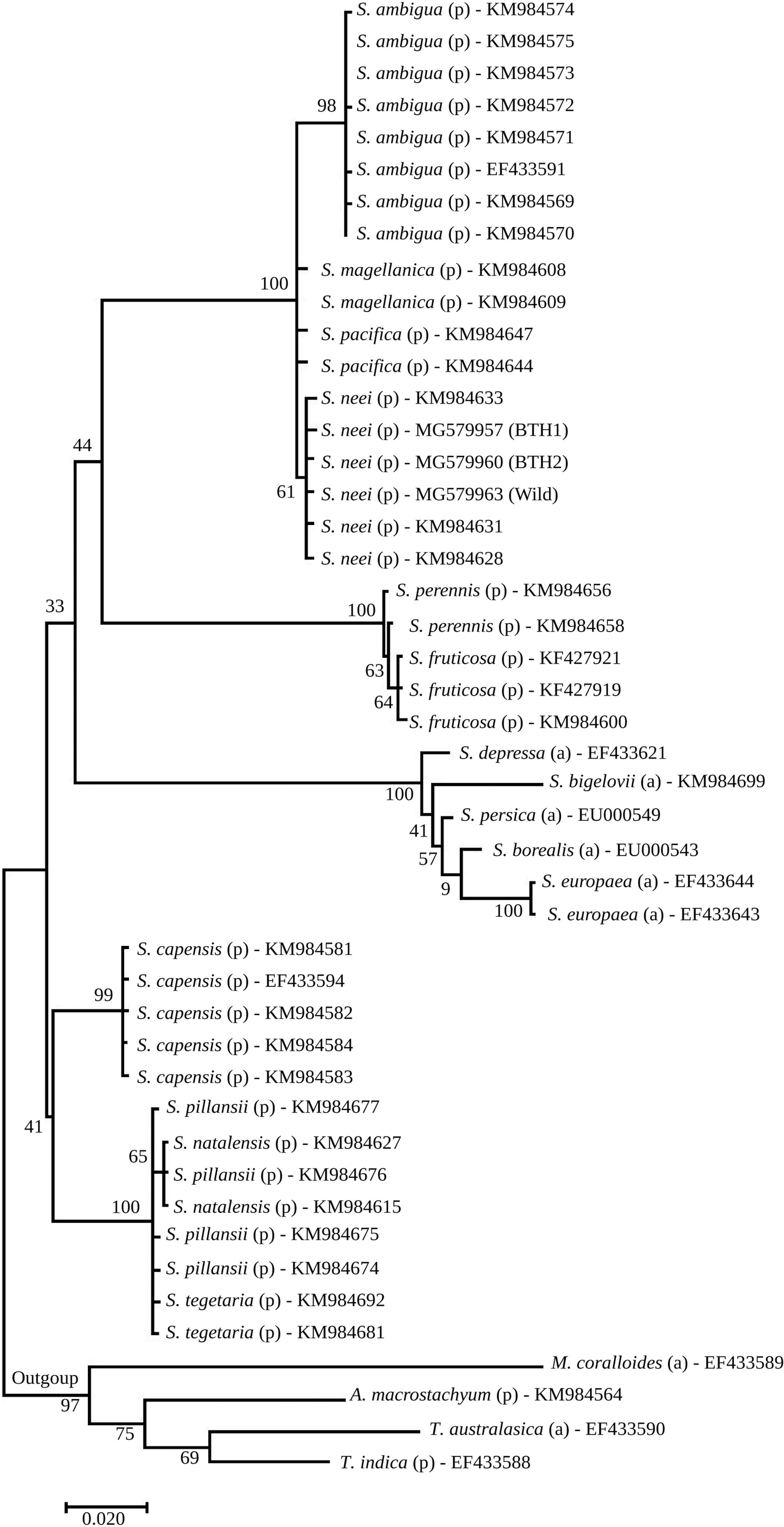
In the ETS marker two mutations were found among the 503 aligned positions of southern Brazilian sequences. Accessions differed at nucleotide positions 199 and 239; BTH1 and BTH2 differed in one nucleotide and they had, respectively, 2 and 1 polymorphic nucleotides in relation to the wild accession (Tab. 2). ETS marker allows us to confirm the identity of Brazilian accessions as S. neei . The BLAST search in GenBank showed that accession KM984631 from Paraguay Charco or accession KM984633 from the margins of Raqui river estuary (south-central Chilean coast) are the closest matches (Fig. 1; Tab. 1).
Maximum likelihood tree based on 46 ETS sequences of Salicornia species; 3 ETS sequences are from accessions of southern Brazil obtained in this study (Wild, BTH1 and BTH2) and 43 ETS sequences from the NCBI database were included. Numbers near nodes are bootstrap values (in percentages) for the internal branches of the tree. The letters “a” and “p” indicate the annual and perennial habits of the species, respectively.
The maximum likelihood tree of ETS sequences indicates that the coastal species of the Salicornia genus in Americas are mainly divided in two major sister clades showing a geographical signal in the phylogenetic relationships of the American species: East coast of North America clade with S. ambigua, and West-East coasts of South American clade with S. neei, S. pacifica and S. magellanica . Analysis of ETS sequences clearly distinguishes (bootstrap percentage = 100) between S. ambigua and S. neei/S. pacifica / S. magellanica clades at the molecular level (Fig. 1).
Discussion
Molecular markers confirm the identity of southern Brazilian accessions as S. neei and show (ETS) small differences among their nucleotides sequences. Previously ITS showed limited variability for other Salicornia species (Kadereit et al. 2006Kadereit G, Mucina L & Freitag H (2006) Phylogeny of Salicornioideae (Chenopodiaceae): diversification, biogeography, and evolutionary trends in leaf and flower morphology. Taxon 55: 617-642. DOI: 10.2307/25065639
https://doi.org/10.2307/25065639...
; de la Fuente et al. 2013)de la Fuente V, Oggerin M, Rufo L, Rodríguez N, Ortuñez E, Sánchez-Mata D & Amilsb R (2013) A micromorphological and phylogenetic study of Sarcocornia A.J.Scott (Chenopodiaceae) on the Iberian Peninsula. Plant Biosystems-An International Journal Dealing with all Aspects of Plant Biology 147: 158-173. DOI: 10.1080/11263504.2012.752414
https://doi.org/10.1080/11263504.2012.75...
, and Steffen et al. (2015)Steffen S, Ball P, Mucina L & Kadereit G (2015) Phylogeny, biogeography and ecological diversification of Sarcocornia (Salicornioideae, Amaranthaceae). Annals of Botany 115: 353-368. DOI: 10.1093/aob/mcu260
https://doi.org/10.1093/aob/mcu260...
concluded that atpB - rbcL spacer data were mostly uninformative among closely related species of Salicornia . Nucleotide substitutions on southern Brazilian Salicornia accessions support the genetic base of prostrate and decumbent growths in pure lineages of local ecomorphotypes. It contrasts to accessions of erect and prostrate forms of Salicornia perennans from NW Kazakhstan that showed no differences between their ETS sequences (Kadereit et al . 2007)Kadereit G, Ball P, Beer S, Mucina L, Sokoloff D, Teege P, Yaprak AE & Freitag H (2007.) A taxonomic nightmare comes true: phylogeny and biogeography of glassworts (Salicornia L., Chenopodiaceae). Taxon 56: 1143-1170. DOI: 10.2307/25065909
https://doi.org/10.2307/25065909...
. However, Doncato & Costa (2018)Doncato KB & Costa CSB (2018) Growth and mineral composition of two lineages of the sea asparagus Sarcocornia ambigua irrigated with shrimp farm saline effluent. Experimental Agriculture 54: 399-416. DOI: 10.1017/S0014479717000096
https://doi.org/10.1017/S001447971700009...
showed that BTH1 and BTH2 sea asparagus lineages had consistent differences in biometrics, production and mineral composition of their biomass under field trials, which were maintained between consecutive progenies of their plants. The analysis of AFLP variation suggested that intraspecific ecotypes might originate repeatedly among some tetraploid species of European Salicornia in adaptation to their specific environments (Teege et al. 2011Teege P, Kadereit JW & Kadereit G (2011) Tetraploid European Salicornia species are best interpreted as ecotypes of multiple origin. Flora 206: 910-920. DOI: 10.1016/j.flora.2011.05.009
https://doi.org/10.1016/j.flora.2011.05....
). After their molecular analysis of Salicornia, Steffen et al. (2015)Steffen S, Ball P, Mucina L & Kadereit G (2015) Phylogeny, biogeography and ecological diversification of Sarcocornia (Salicornioideae, Amaranthaceae). Annals of Botany 115: 353-368. DOI: 10.1093/aob/mcu260
https://doi.org/10.1093/aob/mcu260...
hypothesized that locally adapted prostrate/mat-forming and erect/decumbent ecotypes evolved independently due to selective forces that arise in ecological gradients such as salinity, frequency and duration of flooding.
Current records indicate Salicornia species names recognized in South America (Alonso & Crespo 2008Alonso MA & Crespo MB (2008) Taxonomic and nomenclatural notes on South American taxa of Sarcocornia A.J. Scott (Chenopodiaceae). Annales Botanici Fennici 45: 241-254. DOI: 10.5735/085.045.0401
https://doi.org/10.5735/085.045.0401...
; BFG 2018BFG - The Brazil Flora Group (2018) Brazilian Flora 2020: innovation and collaboration to meet Target 1 of the Global Strategy for Plant Conservation (GSPC). Rodriguésia 69: 1513-1527. DOI: 10.1590/2175-7860201869402
https://doi.org/10.1590/2175-78602018694...
; Steffen et al. 2015Steffen S, Ball P, Mucina L & Kadereit G (2015) Phylogeny, biogeography and ecological diversification of Sarcocornia (Salicornioideae, Amaranthaceae). Annals of Botany 115: 353-368. DOI: 10.1093/aob/mcu260
https://doi.org/10.1093/aob/mcu260...
), of which S. neei is not mentioned as having been collected in Brazil, or even at the Atlantic coast further north of the La Salada lagoon (Pedro Luro, Buenos Aires Province, Argentina; 39° S; Steffen et al. 2015Steffen S, Ball P, Mucina L & Kadereit G (2015) Phylogeny, biogeography and ecological diversification of Sarcocornia (Salicornioideae, Amaranthaceae). Annals of Botany 115: 353-368. DOI: 10.1093/aob/mcu260
https://doi.org/10.1093/aob/mcu260...
). La Salada lagoon accession (KM984628) also shows highly similar ETS sequences to southern Brazilian accessions (Fig. 1; Tab. 1). Alonso & Crespo (2008)Alonso MA & Crespo MB (2008) Taxonomic and nomenclatural notes on South American taxa of Sarcocornia A.J. Scott (Chenopodiaceae). Annales Botanici Fennici 45: 241-254. DOI: 10.5735/085.045.0401
https://doi.org/10.5735/085.045.0401...
recorded accessions of S. neei in the lowlands of central-south Argentina, reaching the Atlantic coast at Valdés Peninsula (Chubut province; ~42° S). Recently Arce et al. (2016)Arce ME, Bianciotto O, Stronati MS, Yepes MS, Blessio AY & Aras FM (2016) Agriculture and sheep production on Patagonian sabkhas with Sarcocornia neei irrigated with seawater (Chubut - Argentina). In: Khan MA, Boër B, Azturk M, Clüsener-Godt M, Gul B & Breckle S-W (eds.) Sabkha ecosystems Vol. V. The Americas. Springer, Berlin. Pp. 275-285. DOI: 10.1007/978-3-319-27093-7_15
https://doi.org/10.1007/978-3-319-27093-...
reported S. neei as an important feeding resource for sheep rearing in saline steppes and marshlands of Patagonia. Salicornia neei presents a wide morphological variation in many parts of its distribution, which, according to Alonso & Crespo (2008)Alonso MA & Crespo MB (2008) Taxonomic and nomenclatural notes on South American taxa of Sarcocornia A.J. Scott (Chenopodiaceae). Annales Botanici Fennici 45: 241-254. DOI: 10.5735/085.045.0401
https://doi.org/10.5735/085.045.0401...
, explains its separation across many taxa. For instance, decumbent and prostrate ecomorphotypes of S. neei were also reported in populations at the Raqui river estuary (Chile; Centro EULA 2011CENTRO EULA (2011) Diagnóstico y propuesta para la conservación y uso sustentable de los humedales lacustres y urbanos principales de la región del Biobío. Centro de Ciencias Ambientales-Universidad de Concepción, Concepción. 134p.). The present genetic record extends the distribution limit of S. neei to the subtropical coast of Brazil.
The southern Brazilian accessions ofsea asparagus certainly are not close to any of the S. ambigua accessions in GenBank, which are restricted to the Northern Hemisphere. As pointed out by Steffen et al. (2015)Steffen S, Ball P, Mucina L & Kadereit G (2015) Phylogeny, biogeography and ecological diversification of Sarcocornia (Salicornioideae, Amaranthaceae). Annals of Botany 115: 353-368. DOI: 10.1093/aob/mcu260
https://doi.org/10.1093/aob/mcu260...
, American perennial Salicornia (= Sarcocornia) is clearly monophyletic and most likely derived from Eurasian ancestors. The ancestral area of the crown node of the American clade was reconstructed to be located in the Northern Hemisphere (either North America or Eurasia or both). Although S. fruticosa/S. perennis clade accessions were widely distributed, along the entire Mediterranean region and even reaching the surrounds of the Dead Sea and the north coast of Africa, these taxa are not present in South America. Similarly, Kadereit et al. (2007)Kadereit G, Ball P, Beer S, Mucina L, Sokoloff D, Teege P, Yaprak AE & Freitag H (2007.) A taxonomic nightmare comes true: phylogeny and biogeography of glassworts (Salicornia L., Chenopodiaceae). Taxon 56: 1143-1170. DOI: 10.2307/25065909
https://doi.org/10.2307/25065909...
and Piirainen et al. (2017)Piirainen M, Liebisch O & Kadereit G (2017) Phylogeny, biogeography, systematics and taxonomy of Salicornioideae (Amaranthaceae/Chenopodiaceae) - A cosmopolitan, highly specialized hygrohalophyte lineage dating back to the Oligocene. Taxon 66: 109-132. DOI: 10.12705/661.6
https://doi.org/10.12705/661.6...
, in the ETS tree, annual Salicornia is sister to a clade comprising Eurasian and North American perennial Salicornia .
Further sampling of wild plants of sea asparagus and molecular evaluation of these accessions across the entire coast of Brazil and in the northern part of South America are required, in order to clarify the distribution limits of S. ambigua and S. neei in the Atlantic region. These samples should be obtained together with detailed morphological data ( i.e. ,growth habit, size of branches and inflorescence, spikes shape and seed indumentum) in order to support the taxonomic assignment. Also, more variable molecular markers using a next generation sequencing approach is needed to reveal the genetic diversification and phylogenetic relationships of S. neei and its close relatives.
Acknowledgments
This work was supported by the Brazilian National Research Council-CNPq (C.S.B.C.; grant number 573884/2008-0-INCTSAL).
References
- Alonso MA & Crespo MB (2008) Taxonomic and nomenclatural notes on South American taxa of Sarcocornia A.J. Scott (Chenopodiaceae). Annales Botanici Fennici 45: 241-254. DOI: 10.5735/085.045.0401
» https://doi.org/10.5735/085.045.0401 - Alonso MA, Crespo MB & Freitag H (2017) Salicornia cuscoensis (Amaranthaceae/Chenopodiaceae), a new species from Peru (South America). Phytotaxa 319: 254-262. DOI: 10.11646/phytotaxa.319.3.4
» https://doi.org/10.11646/phytotaxa.319.3.4 - Arce ME, Bianciotto O, Stronati MS, Yepes MS, Blessio AY & Aras FM (2016) Agriculture and sheep production on Patagonian sabkhas with Sarcocornia neei irrigated with seawater (Chubut - Argentina). In: Khan MA, Boër B, Azturk M, Clüsener-Godt M, Gul B & Breckle S-W (eds.) Sabkha ecosystems Vol. V. The Americas. Springer, Berlin. Pp. 275-285. DOI: 10.1007/978-3-319-27093-7_15
» https://doi.org/10.1007/978-3-319-27093-7_15 - Bertin RL, Gonzag LV, Borges GSC, Azevedo MS, Maltez HF, Heller M, Micke GA, Tavares LBB & Fett R (2014) Nutrient composition and, identification/quantification of major phenolic compounds in Sarcocornia ambigua (Amaranthaceae) using HPLC-ESI-MS/MS. Food Research International 55: 404-411. DOI:10.1016/j.foodres.2013.11.036
» https://doi.org/0.1016/j.foodres.2013.11.036 - BFG - The Brazil Flora Group (2018) Brazilian Flora 2020: innovation and collaboration to meet Target 1 of the Global Strategy for Plant Conservation (GSPC). Rodriguésia 69: 1513-1527. DOI: 10.1590/2175-7860201869402
» https://doi.org/10.1590/2175-7860201869402 - CENTRO EULA (2011) Diagnóstico y propuesta para la conservación y uso sustentable de los humedales lacustres y urbanos principales de la región del Biobío. Centro de Ciencias Ambientales-Universidad de Concepción, Concepción. 134p.
- Costa CSB, Armstrong R, Detrés Y, Koch EW, Bertiller M, Beeskow A, Neves LS, Tourn GM, Bianciotto OA, Pinedo LB, Blessio AY & San Roman N (2006)Effect of ultraviolet-B radiation on salt marsh vegetation: trends of the genus Salicornia along the Americas. Photochemical and Photobiology 82: 878-886. DOI: 10.1562/2005-10-30-RA-729
» https://doi.org/10.1562/2005-10-30-RA-729 - Costa CSB & Davy AJ (1992) Coastal salt marsh communities of Latin America. In: Seeliger U (ed.) Coastal plant communities of Latin America. Academic Press, New York. Pp. 179-199. DOI: 10.1016/B978-0-08-092567-7.50018-0
» https://doi.org/10.1016/B978-0-08-092567-7.50018-0 - Costa CSB & Herrera OB (2016) Halophytic life in Brazilian salt flats: biodiversity, uses and threats. In: Khan MA, Boër B, Azturk M, Clüsener-Godt M, Gul B & Breckle S-W (eds.) Sabkha ecosystems Vol. V. The Americas. Springer, Berlin. Pp. 11-27. DOI: 10.1007/978-3-319-27093-7_2
» https://doi.org/10.1007/978-3-319-27093-7_2 - Costa CSB, Vicenti JRM, Morón-Villarreyes JA, Caldas S, Cardoso LV, Freitas RF & D’oca MGM (2014) Extraction and characterization of lipids from Sarcocornia ambigua meal: a halophyte biomass produced with shrimp farm effluent irrigation. Anais da Academia Brasileira de Ciências 86: 935-943. DOI: 10.1590/0001-3765201420130022
» https://doi.org/10.1590/0001-3765201420130022 - Davy AJ, Bishop GF, Mossman H, Redondo-Goméz S, Castillo JM, Castellanos EM, Luque T & Figueroa ME (2006) Sarcocornia perennis (Miller) A.J.Scott. Biological Flora of the British Isles, n. 244. Journal of Ecology 94: 1035-1048. DOI: 10.1111/j.1365-2745.2006.01156.x
» https://doi.org/10.1111/j.1365-2745.2006.01156.x - de la Fuente V, Oggerin M, Rufo L, Rodríguez N, Ortuñez E, Sánchez-Mata D & Amilsb R (2013) A micromorphological and phylogenetic study of Sarcocornia A.J.Scott (Chenopodiaceae) on the Iberian Peninsula. Plant Biosystems-An International Journal Dealing with all Aspects of Plant Biology 147: 158-173. DOI: 10.1080/11263504.2012.752414
» https://doi.org/10.1080/11263504.2012.752414 - D’oca MGM, Moron-Villarreyes JA, Lemoes JS & Costa CSB (2012) Fatty acids composition in seeds of the South American glasswort Sarcocornia ambigua Anais da Academia Brasileira de Ciências 84: 865-870. DOI: 10.1590/S0001-37652012005000044
» https://doi.org/10.1590/S0001-37652012005000044 - Doncato KB & Costa CSB (2018) Growth and mineral composition of two lineages of the sea asparagus Sarcocornia ambigua irrigated with shrimp farm saline effluent. Experimental Agriculture 54: 399-416. DOI: 10.1017/S0014479717000096
» https://doi.org/10.1017/S0014479717000096 - Doyle JJ (1991) DNA protocols for plants. In: Hewitt G, Johnson AWB & Young JPW (eds.) Molecular techniques in taxonomy. NATO ASI Series H, Cell Biology. Vol. 57. Springer-Verlag, Berlin. Pp. 283-293. DOI: 0.1007/978-3-642-83962-7_18
» https://doi.org/0.1007/978-3-642-83962-7_18 - Freitas RF & Costa CSB (2014) Germination responses to salt stress of two intertidal populations of the perennial glasswort Sarcocornia ambigua Aquatic Botany 117: 12-17. DOI: 10.1016/j.aquabot.2014.04.002
» https://doi.org/10.1016/j.aquabot.2014.04.002 - Isacch JP, Costa CSB, Rodríguez-Gallegol L, Conde D, Escapa M, Gagliardini DA & Iribarne O (2006)Distribution of salt marshes plant communities associated with environmental factors along a latitudinal gradient on the south-east Atlantic coast. Journal of Biogeography 33: 888-900. DOI: 10.1111/j.1365-2699.2006.01461.x
» https://doi.org/10.1111/j.1365-2699.2006.01461.x - Kadereit G, Ball P, Beer S, Mucina L, Sokoloff D, Teege P, Yaprak AE & Freitag H (2007.) A taxonomic nightmare comes true: phylogeny and biogeography of glassworts (Salicornia L., Chenopodiaceae). Taxon 56: 1143-1170. DOI: 10.2307/25065909
» https://doi.org/10.2307/25065909 - Kadereit G, Mucina L & Freitag H (2006) Phylogeny of Salicornioideae (Chenopodiaceae): diversification, biogeography, and evolutionary trends in leaf and flower morphology. Taxon 55: 617-642. DOI: 10.2307/25065639
» https://doi.org/10.2307/25065639 - Kadereit G, Piirainen M, Lambinon J & Van Der Poorten A (2012) Cryptic taxa should have names. Reflections on the glasswort genus Salicornia (Amaranthaceae). Taxon 61: 1227-1239.
- Kumar S, Stecher G & Tamura K (2016) MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Molecular Biology and Evolution 33: 1870-1874. DOI: 10.1093/molbev/msw074
» https://doi.org/10.1093/molbev/msw074 - Piirainen M, Liebisch O & Kadereit G (2017) Phylogeny, biogeography, systematics and taxonomy of Salicornioideae (Amaranthaceae/Chenopodiaceae) - A cosmopolitan, highly specialized hygrohalophyte lineage dating back to the Oligocene. Taxon 66: 109-132. DOI: 10.12705/661.6
» https://doi.org/10.12705/661.6 - Reitz PR (1961) Vegetação da zona marítima de Santa Catarina. Sellowia 13: 17-115.
- Scott AJ (1977) Reinstatement and revision of Salicorniaceae J.Agardh (Caryophyllales). Botanical Journal of the Linnean Society 75: 255-307. DOI: 10.1111/j.1095-8339.1977.tb01493.x
» https://doi.org/10.1111/j.1095-8339.1977.tb01493.x - Steffen S, Ball P, Mucina L & Kadereit G (2015) Phylogeny, biogeography and ecological diversification of Sarcocornia (Salicornioideae, Amaranthaceae). Annals of Botany 115: 353-368. DOI: 10.1093/aob/mcu260
» https://doi.org/10.1093/aob/mcu260 - Teege P, Kadereit JW & Kadereit G (2011) Tetraploid European Salicornia species are best interpreted as ecotypes of multiple origin. Flora 206: 910-920. DOI: 10.1016/j.flora.2011.05.009
» https://doi.org/10.1016/j.flora.2011.05.009 - Ventura Y & Sagi M (2013) Halophyte crop cultivation: the case of Salicornia and Sarcocornia Environmental and Experimental Botany 92:144-153.
- Ventura Y, Eshel A & Pasternak D & Sagi M (2015) The development of halophyte-based agriculture: past and present. Annals of Botany 115:327-331.
Editado por
Datas de Publicação
-
Publicação nesta coleção
08 Ago 2019 -
Data do Fascículo
2019
Histórico
-
Recebido
20 Ago 2017 -
Aceito
08 Jan 2018