Abstracts
The resident population of estuarine dolphin, Sotalia guianensis (P.J. van Bénéden, 1864), in Norte Bay, Santa Catarina, southern Brazil, was studied from 2000 to 2003. Systematic one-day cruises were undertaken aboard a 5 m sail boat equipped with an outboard motor. Applying the focal-group method, information such as geographical position, predominant behavioural patterns, group size, cohesion index and the presence of calves, were registered at every five-minute interval, totalling 198 hours of direct observation of the dolphins. The estuarine dolphin used preferential areas for each behavioural pattern. The mean group size was approximately 29 individuals, and the presence of calves occurred throughout the entire year, but with a significant increase in the warm seasons. The frequency of behavioural patterns and group size varied according to season and time of day, and were strongly associated. The frequency of moving behaviours increased in the cold seasons and along the day. The seasonal variation in the moving behaviours suggest the requirement of a larger area in the cold seasons, probably related to seasonal fluctuations in the abundance of dominant prey items. Higher group sizes were observed while dolphins were foraging. The cohesion index also varied according to the behaviour. Our results showed that group organization and behavioural patterns of this estuarine dolphin population are probably linked to daily and seasonal environment cycles, and also possibly to the condition of being at the furthest southern limit of distribution of the species.
Behaviour; ecology; group organization; Norte Bay; temporal patterns
A população residente de boto-cinza, Sotalia guianensis (P.J. van Bénéden, 1864), da Baía Norte, Santa Catarina, sul do Brasil, foi estudada de 2000 a 2003. Foram realizados cruzeiros sistemáticos de um dia, a bordo de um veleiro de 5 m equipado com motor de popa. Utilizando o método de grupo-focal, informações como posição geográfica, padrão comportamental predominante, tamanho de grupo, índice de coesão e presença de filhotes, foram registradas a cada intervalo de cinco minutos, totalizando 198 horas de observação direta dos botos. O boto-cinza utilizou áreas preferenciais para cada padrão comportamental. O tamanho médio de grupo foi aproximadamente de 29 indivíduos, e a presença de filhotes ocorreu ao longo de todo o ano, porém com aumento significante nas estações quentes. A freqüência dos padrões comportamentais e o tamanho de grupo apresentaram forte correlação, e variaram em relação à estação do ano e hora do dia. Os comportamentos de movimento foram mais freqüentes nas estações frias e ao longo do dia. A variação sazonal destes comportamentos sugere o requerimento de maiores áreas nas estações frias, provavelmente relacionado com as flutuações sazonais de abundância e dominância de presas. Os maiores tamanhos de grupo ocorreram em comportamentos de forrageio. O índice de coesão também variou com o comportamento. Nossos resultados demonstram que a organização de grupo e os padrões comportamentais desta população de boto-cinza são provavelmente influenciados por ciclos ambientais diários e sazonais, e pela condição de limite sul de distribuição para a espécie.
Baía Norte; comportamento; ecologia; organização de grupo; padrões temporais
Seasonal and daily patterns of group size, cohesion and activity of the estuarine dolphin, Sotalia guianensis (P.J. van Bénéden) (Cetacea, Delphinidae), in southern Brazil
Padrões sazonais e diários do tamanho de grupo, coesão e atividade do boto-cinza, Sotalia guianensis (P.J. van Bénéden) (Cetacea, Delphinidae), no sul do Brasil
Fábio G. Daura-Jorge; Leonardo L. Wedekin; Vítor de Q. Piacentini; Paulo C. Simões-Lopes
Departamento de Ecologia e Zoologia, Centro de Ciências Biológicas, Universidade Federal de Santa Catarina, 88040-970, Florianópolis, Santa Catarina, Brasil. E-mail: daurajorge@yahoo.com.br
ABSTRACT
The resident population of estuarine dolphin, Sotalia guianensis (P.J. van Bénéden, 1864), in Norte Bay, Santa Catarina, southern Brazil, was studied from 2000 to 2003. Systematic one-day cruises were undertaken aboard a 5 m sail boat equipped with an outboard motor. Applying the focal-group method, information such as geographical position, predominant behavioural patterns, group size, cohesion index and the presence of calves, were registered at every five-minute interval, totalling 198 hours of direct observation of the dolphins. The estuarine dolphin used preferential areas for each behavioural pattern. The mean group size was approximately 29 individuals, and the presence of calves occurred throughout the entire year, but with a significant increase in the warm seasons. The frequency of behavioural patterns and group size varied according to season and time of day, and were strongly associated. The frequency of moving behaviours increased in the cold seasons and along the day. The seasonal variation in the moving behaviours suggest the requirement of a larger area in the cold seasons, probably related to seasonal fluctuations in the abundance of dominant prey items. Higher group sizes were observed while dolphins were foraging. The cohesion index also varied according to the behaviour. Our results showed that group organization and behavioural patterns of this estuarine dolphin population are probably linked to daily and seasonal environment cycles, and also possibly to the condition of being at the furthest southern limit of distribution of the species.
Key words: Behaviour, ecology, group organization, Norte Bay, temporal patterns.
RESUMO
A população residente de boto-cinza, Sotalia guianensis (P.J. van Bénéden, 1864), da Baía Norte, Santa Catarina, sul do Brasil, foi estudada de 2000 a 2003. Foram realizados cruzeiros sistemáticos de um dia, a bordo de um veleiro de 5 m equipado com motor de popa. Utilizando o método de grupo-focal, informações como posição geográfica, padrão comportamental predominante, tamanho de grupo, índice de coesão e presença de filhotes, foram registradas a cada intervalo de cinco minutos, totalizando 198 horas de observação direta dos botos. O boto-cinza utilizou áreas preferenciais para cada padrão comportamental. O tamanho médio de grupo foi aproximadamente de 29 indivíduos, e a presença de filhotes ocorreu ao longo de todo o ano, porém com aumento significante nas estações quentes. A freqüência dos padrões comportamentais e o tamanho de grupo apresentaram forte correlação, e variaram em relação à estação do ano e hora do dia. Os comportamentos de movimento foram mais freqüentes nas estações frias e ao longo do dia. A variação sazonal destes comportamentos sugere o requerimento de maiores áreas nas estações frias, provavelmente relacionado com as flutuações sazonais de abundância e dominância de presas. Os maiores tamanhos de grupo ocorreram em comportamentos de forrageio. O índice de coesão também variou com o comportamento. Nossos resultados demonstram que a organização de grupo e os padrões comportamentais desta população de boto-cinza são provavelmente influenciados por ciclos ambientais diários e sazonais, e pela condição de limite sul de distribuição para a espécie.
Palavras chave: Baía Norte, comportamento, ecologia, organização de grupo, padrões temporais.
The behaviour of an animal species is influenced by environmental factors that may affect the structure and organization of a population (WILSON 2000) and control both the selection and the way that a habitat is used (KREBS 2001). Understanding this behavioural basis is essential to the knowledge of the ecology of a population and its conservation (SUTHERLAND 1996, CARO 1998).
The link between behavioural and ecological aspects, such as distribution and seasonal abundance of resources, the presence of other species, habitat structure and others, has been addressed in several studies with small cetaceans (WÜRSIG & WÜRSIG 1979, 1980, WELLS et al. 1980, SHANE et al. 1986, SHANE 1990, 1995, GEISE et al. 1999). However, until now, very little is known about the estuarine dolphin, Sotalia guianensis (P.J. van Bénéden, 1864), and this species is classified as "Data Deficient" by the IUCN Red Data Book (IBAMA 2003).
The estuarine dolphin has a wide distribution along the Atlantic coast of Central and South America (BOROBIA et al. 1991). Norte Bay, Santa Catarina, southern Brazil, is the southernmost limit of distribution of the estuarine dolphin (SIMÕES-LOPES 1988). A resident population with a high degree of site fidelity and residency can be found throughout the entire year in the waters of Norte Bay (FLORES 1999). In contrast with the ecological importance of the dolphins, the pressure caused by humans over this population demanded immediate conservation measures in the area, and resulted in the creation of the Environmental Protection Area (EPA) of Anhatomirim in 1992. Recent studies developed in Norte Bay dealt with general aspects of the species ecology (FLORES 1999, FLORES & BAZZALO 2004), spatial use (WEDEKIN et al. in press.), interaction with other species (WEDEKIN et al. 2004) and movement patterns (DAURA-JORGE et al. 2004).
The present work aims at clarifying the behavioural and basic social organization aspects of the estuarine dolphin in Norte Bay and the temporal variations in these parameters. The relationship among behavioural patterns, group size and group spatial cohesion was investigated, as well as how these factors can represent responses to a better adjustment of the dolphins to its environment.
MATERIAL AND METHODS
Study area
Norte Bay is a protected coastal bay that is located on the central coast of Santa Catarina state, adjacent to the Santa Catarina Island, southern Brazil (27º30'S, 48º32'W). It covers an area of approximately 250 km2.
The oceanographic parameters of the water of Norte Bay are very similar to those of the adjacent open waters, which have a strong seasonal variation of characteristics such as water temperature. The mean water temperature on the surface during the cold season is approximately 20ºC, and in the hot season it is approximately 26ºC. The mean depth of the bay is 3.5 m (± 2.37 m), reaching its maximum in both channels (South channel = 22 m, and North channel = 11 m) (DAURA-JORGE et al. 2004).
Data collection
Systematic one-day cruises (daily effort of about 10 h) were undertaken aboard a 5 m sail boat equipped with an outboard motor. The study area was surveyed counter-clockwise, leaving from the eastern coast of the bay. Additional and opportunistic routes were undertaken in order to cover the area homogeneously. After a group of dolphin was encountered, the route was interrupted and the dolphins were followed using the focal-group method (LEHNER 1996). Data such as geographic position (GPS Garmin), predominant behavioural pattern, group size, cohesion index and the presence of calves, were registered at every five-minute interval.
Following LEHNER (1996), behaviours may be regarded as two distinct units: events, which are instantaneous displays; and states (denoted here as behavioural pattern, or simply behaviour), which have appreciable duration and were the focus of all analyses carried out in this article. The predominant behavioural pattern for each interval was defined based on the events and other evidence observed, which included aerial and surface activities, the presence of seabirds, and swimming speed and direction (MANN 1999).
The behavioural patterns considered in the present work were adapted from SHANE (1990), as follow: feeding (F) short and asynchronised dives, abundant surface events, movements in varying directions and, many times, presence of seabirds; travelling (T) moving steadily in one direction, with synchronised dives; travel/feeding (Tf) behavioural characteristics similar to the feeding pattern, but moving in a defined direction, similar to travelling; resting (R) lack of surface events, slow movements and undefined direction; and socializing (S) vigorous and isolated interaction activities among the individuals while lacking evidence of any other predominant behavioural pattern of the group.
A group was defined as any aggregation of dolphins observed in an apparent association, frequently, but not always, engaged in the same activity, in which it was possible to directly count the number of individuals and to determine the predominant group activity (modified from SHANE 1990). The counts of individuals lasted approximately 30 seconds and were done at the moment with the best view of the group during the five-minute interval. The group had a very dynamic organization and smaller social unit (called subgroups), composed of few individuals, were also possible to identify, but they were considered to be part of the group.
For the analysis of spatial cohesion of the group, we used a cohesion index (CI). Three arbitrary degrees of spatial dispersion of the group were established: low cohesion (CI 1) dolphins were scattered through a sometimes large area, more than 100 m wide; intermediate cohesion (CI 2) dolphins occupied an area between approximately 50 and 100 m wide; and high cohesion (CI 3) dolphins were highly aggregated occupying a small area, usually smaller than 30 m.
The counting of individuals, the determination of the behaviour and the cohesion index were performed by experienced researchers, who were always the same in the whole period of study, reducing possible inter-observer bias (see LEHNER 1996).
Data analysis
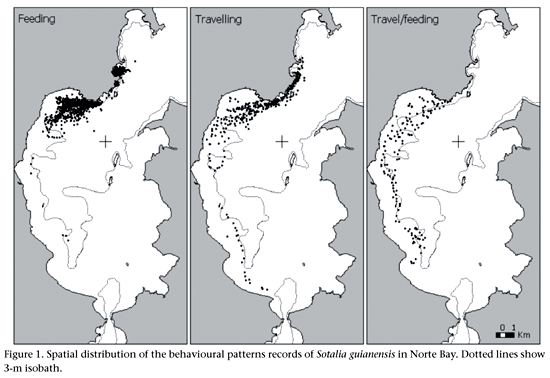
The geographic positions of each behavioural pattern were plotted in a digitised nautical chart of Norte Bay using ArcView GIS 3.2 (Environment Systems Research Institute ESRI). As a result, we generated maps of distribution for each behavioural pattern to verify if the estuarine dolphins used preferential areas for different activities.
For all seasonal variation analyses we used only data from Spring 2001 to Winter 2002, which were the four consecutive seasons with the most regular sampling effort throughout the study period. The whole dataset was used in the other analyses. The resting and socializing states were excluded from the analyses of behaviours patterns due to their occasional occurrence during the observations. For the analysis of the relationship between behavioural pattern and seasonal and daily variations of group size, and only when supported by previous statistical analyses, we merged feeding and travel/feeding into a single functional category called foraging.
The day was divided in three periods of equal duration for the analyses of daily variation of group size and behaviour: 1) 8:00-10:59 h; 2) 11:00-13:59 h; 3) 14:00-17:00 h. Data gathered before 8:00 and after 17:00 h, due to a small sample size, were excluded from the analyses.
The seasonal and daily variations of the frequency of behavioural patterns, and the association of the cohesion index with behaviour were tested using the Independency Chi-Square test (SOKAL & ROHLF 1995). To assess the seasonal and daily variation of group size, and if group size varied among behavioural patterns, we used the Kruskal-Wallis test (SOKAL & ROHLF 1995). The association between group size and frequency of behavioural pattern was measured with Pearson Correlation test, and, lastly, to verify if the absolute frequency of calves varied seasonally we also used the Independency Chi-Square test (SOKAL & ROHLF 1995). All analyses were conducted using the Statistica 5.0 software.
RESULTS
Field effort
Between July 2000 and December 2002, 69 one-day cruises were made to study cetaceans in Norte Bay, totalling 422 hours of sampling effort, of which 198 hours (46.4% of the effort) corresponded to direct observation of the dolphins. The continuous observation time of the same group of dolphins varied from 0.58 h to 8.08 h, and the mean duration was 4.31 h. In 29% of the sampled days groups of dolphins were not encountered. We never observed more than one group of dolphins in one day of sampling effort, even after leaving the first encountered group of dolphins after some hours of observation, and sampling other areas. The sampling characteristics of the most regular consecutive four seasons, which were used in the seasonal analyses, are summarized in table I.
Spatial distribution of behaviour
The dolphins used distinct areas for each behavioural pattern (Fig. 1). Feeding occurred preferentially in two distinct areas of Norte Bay: São Miguel and Currais bays. The area used for travelling and travel/feeding overlapped extensively, and were characterized by a large stretch parallel to the coast and at the same depth range. Resting and socializing were restricted to Currais Bay and its surroundings.
Seasonal and daily variations of behaviour
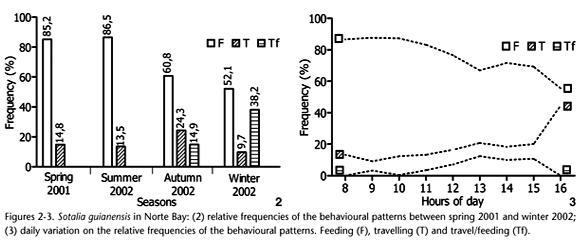
The frequency of behavioural patterns varied among different seasons of the year (c2 = 134.3088; df = 6; p < 0.05). Feeding was the most frequent behaviour observed over the entire study period, but tended to decrease in the cold seasons (Autumn and Winter 2002), while travel/feeding and travelling increased substantially (Fig. 2). Resting and socializing were rarely observed (2.58% and 0.55%, respectively).
The frequency of behavioural patterns varied among different periods of day (c2 = 44.5379; df = 4; p < 0.05). There was an inverse trend along the day between moving behaviours (travelling and travel/feeding) and feeding (Fig. 3). While the frequency of feeding decreased progressively, travelling and travel/feeding tended to increase in frequency.
Seasonal and daily variations of group size
The mean group size of Sotalia guianensis in the study area was 29 individuals (Sd = 9.8; range = 1-59). The most observed value (mode) was 29 individuals and small groups of 1 to 10 individuals were rare (4.2 %). The small groups of dolphins were never observed far from a larger aggregation of individuals (more than 1 km apart from each other).
Group size varied along the seasons of the year (Kruskal-Wallis: H = 112.839; df = 3/1241; p < 0.05; Fig. 4). The Spring 2001 presented the lowest mean group size (25.5 individuals), which increased in Summer and Autumn 2002 (30.2 and 34.0 individuals, respectively) and decreased again in Winter 2002 (31.86 individuals).
The mean group size varied along the day (Kruskal-Wallis: H = 72.197; df = 2/2039; p < 0.05). The largest mean group size was observed in the beginning of the day (mean: 31.1 individuals) and decreased progressively until late afternoon (mean: 25.6 individuals; Fig. 5).
Relationship between group size and behaviour
Group size varied among different behavioural patterns (Kruskal-Wallis: H = 223.608; df = 2/2101; p < 0.05). Group size of the estuarine dolphin when feeding and travel/feeding was similar (mean = 30.4 and 30.6 individuals, respectively) and higher than during travelling (mean = 21.7 individuals; Fig. 6), thus supporting our merging of feeding and travel/feeding states into a category named "foraging".
There was a strong positive association of group size and frequency of foraging (feeding + travel/feeding) through the seasons (r = 0.86; n = 8; p < 0.05) and a moderate positive association between these two parameters throughout the day (r = 0.78; n = 11; p < 0.05).
Group spatial cohesion and calves presence
Spatial cohesion of estuarine dolphin groups, similarly as group size, varied regarding the behavioural pattern. There was a significant variation of cohesion index among the behavioural patterns (c2 = 176.402; df = 4; p < 0.05). The higher degree of group spatial cohesion was observed while dolphins were feeding, and the lowest degree was observed when dolphins were travel/feeding (Fig. 7).
In every group we followed during the study period we observed the presence of at least one calf. The seasonal difference in the frequency of records of one or more calves in the group could potentially reflect a season of births. A total of 613 records (24.7%) were obtained of at least one calf presence in the group. In the warm seasons there was a higher frequency of records of calves in groups of dolphins (Spring 2001 = 30.5% and Summer 2002 = 29.7%) compared to the cold seasons (Autumn 2002 = 24% and Winter 2002 = 19.8%), and this variation was statistically significant (c2 = 9.258; df = 3; p < 0.05).
DISCUSSION
The predominance of feeding in more sheltered and/or lower water circulation areas, such as Currais and São Miguel bays, as well as an overlap of movement behaviour with the 3 m isobath, as mentioned earlier by FLORES & BAZZALO (2004), demonstrate that different areas were used for distinct behaviours of the estuarine dolphin in Norte Bay. This reflects the habitat heterogeneity of the coastal waters of Norte Bay, requiring distinct behavioural responses by the dolphins. The choice of preferential areas for given activities seems to be a common characteristic of estuarine dolphin populations (ARAÚJO et al. 2001, CREMER et al. 2004) as well as other coastal small cetaceans such as Tursiops truncatus (Montagu, 1821) and Sousa chinensis (Osbeck, 1765) (SHANE 1980, GRIGG & MARKOWITZ 1997, KARCZMARSKI et al. 2000).
Resting and socializing behaviours of the estuarine dolphin are difficult to be recognized in the field and tend to be recorded less frequently. In the present study these behaviours were probably underestimated. The predominance of large groups in the area and the use of focal-group sampling, focussing on the predominant activities, may mask the occurrence of such behaviours (see MANN 1999).
Feeding was the most frequent behaviour in the entire study. The same tendency was observed by C. BONIN (pers. comm.) for the estuarine dolphin in Guaraqueçaba Bay, southeastern Brazil, however, higher frequencies of travelling activities were observed in Guanabara Bay, eastern Brazil (GEISE et al. 1999), and Babitonga Bay, southern Brazil (M. CREMER, pers. comm.).
The temporal variation of behavioural states points to a higher feeding activity in the warm seasons and a tendency of moving more (travelling and travel/feeding) in the cold seasons. A similar pattern was observed in recent analyses of group home range size (WEDEKIN et al. in press.) and movement intensity (DAURA-JORGE et al. 2004) of this same population. Both studies suggest the need of wider spatial requirements during cold seasons, probably as a consequence of a fluctuation in the abundance of feeding resources.
Feeding activity was the most frequent one over the whole day and tended to decrease in the afternoon, when travelling increased in intensity. This pattern of activity was observed on almost all days sampled. A daily variation of behaviour or the regular cycle of daily activities may be influenced by photoperiod and the tide regime (SHANE et al. 1986), and even wind speed, but this must be an indirect relation, and the dolphins would actually be responding to prey movement. Another factor that could subject this pattern is the resource depletion in a given area, such as food, that would cause the animal to move to other sites looking for the depleted resource (SUTHERLAND 1996). In the case of Norte Bay, correlations between behavioural activities and environmental variables (such as tide and wind) may elucidate the daily pattern found, and further study on the prey dynamics may establish the links among these various components.
The presence of calves occurs throughout an entire year in Norte Bay, but apparently there is a reproductive season. The difference in frequency of calves was significant among seasons with a slight increase in the warm seasons. This finding agrees with the studies of GEISE et al. (1999) and LODI (2003), who found a summit in calves' frequency for the estuarine dolphin in spring and summer. On the other hand, ROSAS & MONTEIRO-FILHO (2002) did not find this seasonal pattern in their study about the reproduction of the estuarine dolphins. In this way, additional studies focused specifically on reproduction would be desirable to clarify this subject concerning the population of Norte Bay.
The seasonal and daily variations of group size were strongly associated with seasonal and daily variations of behavioural patterns. Feeding and travel/feeding presented the highest group sizes in Norte Bay, and the increasing of their frequencies in a season or period of day may have lead to an increase of the mean group size. This increase may be caused by a tendency to higher counts for given behaviours. When a group is too dispersed, the count becomes more difficult and may be underestimated. The feeding behaviour, which presented the most aggregated spatial conformation (lower cohesion index), and the travel/feeding, which presented the most dispersed one, had extremely similar group size, though. Nonetheless, the breathing/diving intervals and the travelling speed of each behaviour may influence the counts, so that the group size variations found may be a consequence of these factors instead of real fluctuations of the number of individuals present in the group.
The Sotalia guianensis population of Norte Bay presents a singular group structure. The mean group size recorded (29 individuals) was higher than what is normally observed for this species in other areas along its distribution (GEISE 1991, OLIVEIRA et al. 1995, GEISE et al. 1999, ARAÚJO et al. 2001, EDWARDS & SCHNELL 2001). Another interesting characteristic is that two or more groups were never registered using different areas in Norte Bay. Instead, a single formation was always observed, with a high mean group size, extremely dynamic but apparently stable, in which almost all (or probably all) individuals were present. This is different from what is observed in other areas of occurrence of the species, where several different groups can be observed within a single sampling day (LW, FDJ and PCSL, pers. obs.). Although we did not show results to confirm this hypothesis, this view is corroborated by studies about the ecology of this population, which showed a high degree of residency and site fidelity (FLORES 1999), extensive overlap among individual home-ranges (FLORES & BAZZALO 2004), and a highly aggregated pattern of spatial usage (WEDEKIN et al. in press.).
Several studies of small cetaceans suggest that the size and conformation of a group may be related to habitat structure and social-environmental aspects of the populations (e.g. NORRIS & DOHL 1980, WÜRSIG & WÜRSIG 1980, WELLS et al. 1987, SHANE 1990, BEL'KOVICH et al. 1991, WÜRSIG et al. 1991, BAIRD & DILL 1996, GYGAX 2002). The potential advantages of living in groups have been of great interest and speculation. According to WILSON (1975), the group size is among the main characteristics of the social organization of a population. The causal relationship between habitat and group size is yet to be known. According to PULLIAN & CARACO (1987), a great abundance and aggregation of food resources and the presence of predators are two characteristics of the environment that would propitiate the formation of large groups as those observed in Norte Bay for the estuarine dolphin. These two issues are still poorly known in Norte Bay and other areas of occurrence of this species. However, the presence of the larger bottlenose dolphin in Norte Bay is also a trait that may influence the social organization and group formation of the estuarine dolphin, since both species have already been observed in an aggressive interspecific interaction in the area (WEDEKIN et al. 2004). It is also important to consider that the population studied here is found in the southernmost limit of distribution of the species. The geographic outliers of a species are likely to occur in unusual, atypical or in marginal and stressful conditions (see review in LESICA & ALLENDORF 1995), and thus we can expect that the behavioural responses of an animal to this conditions may be equally unusual and atypical.
Our results showed that group organization and behavioural patterns of this estuarine dolphin population are probably linked to daily and seasonal environment cycles, and also possibly to the condition of being at the furthest limit of distribution of the species. Behaving in different manners along the temporal and spatial axis are forms of better exploring and adapting to the varying environment. We leave an open road for future research projects aiming at test and explore how specifically the biotic and abiotic factors of the environment condition these behavioural responses, although we already proposed some hypothesis based on the available data.
ACKNOWLEDGMENTS
The Brazilian Research Bureau (Conselho Nacional de Desenvolvimento Científico e Tecnológico CNPq) funded P.C. Simões-Lopes (Proc. 302749/2002-0) and F.G. Daura-Jorge (BCI), Socioambiental Consultores Associados Ltda. funded L.L. Wedekin; Mercury do Brasil provided an outboard motor. We also thank Marta Cremer, Emygdio Monteiro-Filho, Maurício Graipel and Karim H. Lüchmann for their contributions and suggestions.
Received in 06.IV.2005; accepted in 05.XI.2005.
- ARAÚJO, J.P.; J.Z.O. PASSAVANTE & A.S. SOUTO. 2001. Behavior of the estuarine dolphin, Sotalia guianensis, at Dolphin Bay, Pipa, Rio Grande do Norte, Brazil. Tropical Oceanography, Recife, 29: 13-23.
- BAIRD, R.W. & L.M. DILL. 1996. Ecological and social determinants of group size in transient killer whales. Behavioral Ecology, Oxford, 7: 408-416.
- BEL'KOVICH, V.M.; A.V. AGAFONOV; O.V. YEFREMENKOVA; L.B. KOZAROVITSKY & S.P. KHARITONOV. 1991. Herd structure, hunting, and play bottlenose dolphins in Black Sea, p. 17-38. In: K.A. PRYOR & K.S. NORRIS (Eds). Dolphin Societies: discoveries and puzzles. Berkeley, University of California Press, 397p.
- BOROBIA, M.; S. SICILIANO; L. LODI & W. HOEK. 1991. Distribution of the South American dolphin Sotalia fluviatilis Canadian Journal of Zoology, Ottawa, 69: 1025-1039.
- CARO, T. 1998. The significance of behavioral ecology for conservation biology, p. 3-26. In: T. CARO (Ed.). Behavioral ecology and conservation biology. Oxford, Oxford University Press, 480p.
- CREMER, M.J.; F.A.S. HARDT.; A.J.T. JUNIOR.; P.C.A. SIMÕES-LOPES & J.S.R. PIRES. 2004. Core areas changes in Sotalia guianensis (Cetacea, Delphinidae) population in Babitonga Bay, Santa Catarina. Revista Univille, Joinville, 9: 130-134.
- DAURA-JORGE, F.G.; L.L. WEDEKIN & P.C. SIMÕES-LOPES. 2004. Variação sazonal na intensidade dos deslocamentos do boto-cinza, Sotalia guianensis (Cetacea: Delphinidae), na Baía Norte da Ilha de Santa Catarina. Biotemas, Florianópolis, 17: 203-216.
- EDWARDS, H.H. & G.D. SCHNELL. 2001. Status and ecology of Sotalia fluviatilis in the Cayos Miskito Reserve, Nicaragua. Marine Mammal Science, California, 17: 445-472.
- FLORES, P.A.C. 1999. Preliminary results of a photoidentification study of the marine tucuxi, Sotalia fluviatilis, in Southern Brazil. Marine Mammal Science, California, 15: 840-847.
- FLORES, P.A.C. & M. BAZZALO. 2004. Home range and movement patterns of the marine tucuxi dolphin, Sotaia fluviatilis, in Baía Norte, southern Brazil. Latin American Journal of Aquatic Mammals, Rio de Janeiro, 3: 37-52.
- GEISE, L. 1991. Sotalia guianensis (Cetacea, Delphinidae) population in the Guanabara Bay, Rio de Janeiro, Brazil. Mammalia, Paris, 55: 371-379.
- GEISE, L.; N. GOMES & R. CERQUEIRA. 1999. Behavior, habitat use and population size of Sotalia fluviatilis (Gervais, 1853) in the Cananéia estuary region, São Paulo, Brazil. Brazilian Journal of Biology, São Carlos, 59: 183-194.
- GRIGG, E. & H. MARKOWITZ. 1997. Habitat use by bottlenose dolphins (Tursiops truncatus) at Turneffe Atoll, Belize. Aquatic Mammals, Illinois, 23: 163-170.
- GYGAX, L. 2002. Evolution of group size in the dolphins and porpoises: interspecific consistency of intraspecific patterns. Behavioral Ecology, Oxford, 13: 583-590.
- IBAMA. 2003. Mamíferos Aquáticos do Brasil: Plano de Ação Brasília, Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis, 79p.
-
16KARCZMARSKI, L.; V.G. COCKCROFT & A. MCLACHLAN. 2000. Habitat use and preferences of Indo-pacific humpback dolphins Sousa chinensis in Algoa Bay, South Africa. Marine Mammal Science, California, 16: 65-79.
- KREBS, C. J. 2001. Ecology: the experimental analysis of distribution and abundance. San Francisco, Benjamin Cummings, 695p.
- LEHNER, P.N. 1996. Handbook of ethological methods. Cambridge, University Press, 672p.
- LESICA, P. & F.W. ALLENDORF. 1995. When are peripheral populations valuable for conservation? Conservation Biology, Florida, 9: 753-760.
- LODI, L. 2003. Tamanho e composição de grupo dos botos-cinza, Sotalia guianensis (van Bénéden, 1864) (Cetacea, Delphinidae), na Baía de Paraty, Rio de Janeiro, Brasil. Atlântica, Rio Grande, 25: 135-146.
- MANN, J. 1999. Behavioral sampling methods for cetaceans: a review and critique. Marine Mammal Science, California, 15: 102-122.
- NORRIS, K.S. & T.P. DOHL. 1980. The structure and functions of cetacean schools, p. 211-261. In: L.M. Herman (Ed.). Cetacean behavior: mechanisms and functions. New York, John Wiley and Sons, 463p.
- OLIVEIRA, J.A.; F.J.C. ÁVILA; T.T.A. JÚNIOR; M. A.A. FURTADO-NETO; C. MONTEIRO-NETO. 1995. Monitoramento do boto-cinza, Sotalia fluviatilis (Cetacea: Delphinidae) em Fortaleza, Estado do Ceará, Brasil. 1995. Arquivo de Ciências do Mar, Fortaleza, 29: 28-35.
- PULLIAM, R. & T. CARACO. 1987. Living in groups: Is there an optimal group size?, p. 122-147. In: J.R. KREBS & N.B. DAVIES (Eds). Behavioural ecology: an evolutionary approach. Oxford, Oxford University Press, 493p.
- ROSAS, F.C.W. & E.L.A. MONTEIRO-FILHO. 2002. Reproduction of the estuarine dolphin (Sotalia guianensis) on the coast of Paraná, southern Brazil. Journal of Mammalogy, Lawrence, 83: 507-515.
- SHANE, S.H. 1980. Occurrence, movements, and distribution of bottlenose dolphins, Tursiops truncates, in southern Texas. Fishery Bulletin, California, 78: 593-601.
- SHANE, S.H. 1990. Behavior and ecology of the bottlenose dolphins at Sanibel Island, Florida, p. 245-265. In: S. LEATHERWOOD & R.R. REEVES (Eds). The Bottlenose Dolphin. San Diego, Academic Press, 643p.
- SHANE, S.H. 1995. Relationship between pilot whales and Risso's dolphins at Santa Catalina Island, California, USA. Marine Ecology Progress Series, Oldendorf, 123: 5-11.
- SHANE, S.H.; R.S. WELLS & B. WÜRSIG. 1986. Ecology, behavior and social organization of the bottlenose dolphin: a review. Marine Mammal Science, California, 2: 34-63.
- SIMÕES-LOPES, P.C. 1988. Ocorrência de uma população de Sotalia fluviatilis (Gervais), 1853, (Cetacea, Delphinidae) no limite sul de sua distribuição, Santa Catarina, Brasil. Biotemas, Florianópolis, 1: 57-62.
- SOKAL, R.R. & F.J. ROHLF. 1995. Biometry. New York, W.H. Freeman, 887p.
- SUTHERLAND, W.J. 1996. From individual behaviour to population ecology. Oxford, Oxford University Press, 213p.
- WEDEKIN, L.L.; F.G. DAURA-JORGE; V.Q. PIACENTINI; P.C. SIMÕES-LOPES. (in press). Seasonal variation in spatial usage by the estuarine dolphin, Sotalia guianensis (Cetacea: Delphinidae), at its southern limit of distribution. Brazilian Journal of Biology, São Carlos.
- WEDEKIN, L.L.; F.G. DAURA-JORGE & P.C. SIMÕES-LOPES. 2004. An Aggressive Interaction Between Bottlenose Dolphins (Tursiops truncatus) and Estuarine Dolphins (Sotalia guianensis) in Southern Brazil. Aquatic Mammals, Illinois, 30: 391-397.
- WELLS, R.S.; A.B. IRVINE & M.D. SCOTT. 1980. The social ecology of inshore odontocetes, p. 263-317. In: L.M. HERMAN (Ed.). Cetacean behavior, mechanisms and functions. New York, John Wiley, 463p.
- WELLS, R.S.; M.D. SCOTT & A.B. IRVINE. 1987. The social structure of free-ranging bottlenose dolphins, p. 247-305. In: H.H. GENOWAYS (Ed.). Current Mammalogy. New York, Plenum Press, 519p.
- WILSON, D.S. 1975. A theory of group selection. Proceeding of the National Academy of Sciences of the United States of America, Washington, 72: 143-146.
- WILSON, E.O. 2000. Sociobiology: the new synthesis. Cambridge, The Belknap Press of Harvard University Press, 697p.
- WÜRSIG, B. & M. WÜRSIG. 1979. Behavior and ecology of the bottlenose dolphin, Tursiops truncatus, in the South Atlantic. Fishery Bulletin, California, 77: 399-412.
- WÜRSIG, B. & M. WÜRSIG. 1980. Behavior and ecology of the dusky dolphin, Lagenorhynchus obscurus, in the South Atlantic. Fishery Bulletin, California, 77: 871-890.
- WÜRSIG, B.; F. CIPRIANO & M. WÜRSIG. 1991. Information from radio and theodolite tracking studies, p. 79-111. In: K.A. PRYOR & K.S. NORRIS (Eds.). Dolphin Societies: discoveries and puzzles. Berkeley, University of California Press, 397p.
Publication Dates
-
Publication in this collection
09 Mar 2006 -
Date of issue
Dec 2005
History
-
Received
06 Apr 2005 -
Accepted
05 Nov 2005