Abstracts
Understanding of the morphogenetic characteristics allows comprehension of aspects related to the form and function of forage plants, providing opportunities to identify potentially high productive plants and distinct defoliation management requirements. The objective of this experiment was to carry out a comparative study of ten tropical forage grasses using morphogenetic and structural variables. Treatments corresponded to grasses of the Panicum genus (P. maximum cultivars Tanzânia and Mombaça) and the Brachiaria genus (B. brizantha cultivars Piatã, Marandu, Xaraés, Capiporã and Arapoty; B. decumbens cultivar Basilisk; B. humidicola cultivars Comum and Tupi), evaluated under free growth conditions.. Response variables were leaf appearance and elongation rates, phyllochron, stem elongation rate, final leaf length, number of live leaves per tiller, leaf life span and tiller appearance, death and survival rates. There was difference between cultivars for these variables, indicating a large variability within plants. Considering the morphogenetic and structural variables of leaves and tillers, B. brizantha had a similar pattern of variation to those of P. maximum, and B. Decumbens, similar to those of B. humidicola. Tiller appearance was large at the onset of the experiment and second and third generations comprised the large majority of tiller population for the cultivars Mombaça, Tanzânia, Xaraés and Capiporã. Group analysis based on plant morphogenetic and structural characteristics of leaves and tillers was an interesting way of grouping cultivars, indicating the importance and the potential of using morphogenesis in plant selection and evaluation programmes.
Brachiaria; Panicum; morphogenetic and structural variables
O estudo das características morfogênicas permite compreender aspectos relativos à forma e função das plantas forrageiras além de possibilitar identificar plantas com potenciais produtivos e exigências de manejo diferentes. O objetivo deste trabalho foi avaliar, comparativamente, dez plantas forrageiras tropicais por meio de variáveis morfogênicas e estruturais. Os tratamentos corresponderam a cultivares pertencentes ao gênero Panicum (P. maximum cvs. Tanzânia e Mombaça) e ao gênero Brachiaria (B. brizantha cvs. Piatã, Marandu, Xaraés, Capiporã, e Arapoty; B. decumbens cv. Basilisk; B. humidicola cvs. Comum e Tupi), avaliados sob condições de crescimento livre. Foram avaliadas a taxa de aparecimento e alongamento de folhas, filocrono, taxa de alongamento de colmos, comprimento final da lâmina foliar, número de folhas vivas por perfilho, duração de vida das folhas e padrão demográfico de perfilhamento. Houve diferença entre cultivares para as variáveis, indicando grande variabilidade no material estudado. Considerando as características morfogênicas e estruturais de folhas e perfilhos, as B. brizantha apresentaram comportamento parecido com os P. maximum, e B. decumbens mais próximo das B. humidicola. Verificou-se que o aparecimento de perfilhos foi inicialmente elevado e que a segunda e, ou, terceira gerações contribuíram de forma significativa para o número total de perfilhos dos cultivares Mombaça, Tanzânia, Xaraés e Capiporã. A análise de agrupamento baseada nas características morfogênicas e estruturais de folhas e perfilhos foi um método interessante para agrupar cultivares, demonstrando a importância e o potencial de uso da morfogênese em trabalhos de seleção e avaliação de plantas forrageiras.
Brachiaria; Panicum; variáveis morfogênicas e estruturais
ANIMAL SCIENCE AND PASTURES
Morphogenetic and structural comparative characterization of tropical forage grass cultivars under free growth
Caracterização morfogênica e estrutural comparativa de cultivares de gramíneas forrageiras tropicais sob crescimento livre
Márcia Cristina Teixeira da SilveiraV; Domicio do Nascimento JúniorI,* * Corresponding author < domicio@ufv.br> ; Sila Carneiro da SilvaII; Valéria Pacheco Batista EuclidesIII; Denise Bataglin MontagnerIII; André Fischer SbrissiaIV; Carlindo Santos RodriguesV; Bráulio Maia de Lana SousaV; Karine da Silva PenaV; Hélio Henrique VilelaV
IUFV Depto. de Zootecnia, Av. P.H. Rolfs, s/n Campus Universitário 36570-000 Viçosa, MG Brasil
IIUSP/ESALQ Depto. de Zootecnia, Av. Pádua Dias, n.11 13418-900 Piracicaba, SP Brasil
IIIEmbrapa Gado de Corte, BR 262, km 4 C.P. 154 - 79002-970 Campo Grande, MS Brasil
IVUDESC Depto. de Zootecnia, Rua Luiz de Camões, n 2090 Centro Agroveterinário - 88520-000 Lages, SC Brasil
VUFV Programa de Pós-Graduação em Zootecnia
ABSTRACT
Understanding of the morphogenetic characteristics allows comprehension of aspects related to the form and function of forage plants, providing opportunities to identify potentially high productive plants and distinct defoliation management requirements. The objective of this experiment was to carry out a comparative study of ten tropical forage grasses using morphogenetic and structural variables. Treatments corresponded to grasses of the Panicum genus (P. maximum cultivars Tanzânia and Mombaça) and the Brachiaria genus (B. brizantha cultivars Piatã, Marandu, Xaraés, Capiporã and Arapoty; B. decumbens cultivar Basilisk; B. humidicola cultivars Comum and Tupi), evaluated under free growth conditions.. Response variables were leaf appearance and elongation rates, phyllochron, stem elongation rate, final leaf length, number of live leaves per tiller, leaf life span and tiller appearance, death and survival rates. There was difference between cultivars for these variables, indicating a large variability within plants. Considering the morphogenetic and structural variables of leaves and tillers, B. brizantha had a similar pattern of variation to those of P. maximum, and B. Decumbens, similar to those of B. humidicola. Tiller appearance was large at the onset of the experiment and second and third generations comprised the large majority of tiller population for the cultivars Mombaça, Tanzânia, Xaraés and Capiporã. Group analysis based on plant morphogenetic and structural characteristics of leaves and tillers was an interesting way of grouping cultivars, indicating the importance and the potential of using morphogenesis in plant selection and evaluation programmes.
Key words:Brachiaria, Panicum, morphogenetic and structural variables
RESUMO
O estudo das características morfogênicas permite compreender aspectos relativos à forma e função das plantas forrageiras além de possibilitar identificar plantas com potenciais produtivos e exigências de manejo diferentes. O objetivo deste trabalho foi avaliar, comparativamente, dez plantas forrageiras tropicais por meio de variáveis morfogênicas e estruturais. Os tratamentos corresponderam a cultivares pertencentes ao gênero Panicum (P. maximum cvs. Tanzânia e Mombaça) e ao gênero Brachiaria (B. brizantha cvs. Piatã, Marandu, Xaraés, Capiporã, e Arapoty; B. decumbens cv. Basilisk; B. humidicola cvs. Comum e Tupi), avaliados sob condições de crescimento livre. Foram avaliadas a taxa de aparecimento e alongamento de folhas, filocrono, taxa de alongamento de colmos, comprimento final da lâmina foliar, número de folhas vivas por perfilho, duração de vida das folhas e padrão demográfico de perfilhamento. Houve diferença entre cultivares para as variáveis, indicando grande variabilidade no material estudado. Considerando as características morfogênicas e estruturais de folhas e perfilhos, as B. brizantha apresentaram comportamento parecido com os P. maximum, e B. decumbens mais próximo das B. humidicola. Verificou-se que o aparecimento de perfilhos foi inicialmente elevado e que a segunda e, ou, terceira gerações contribuíram de forma significativa para o número total de perfilhos dos cultivares Mombaça, Tanzânia, Xaraés e Capiporã. A análise de agrupamento baseada nas características morfogênicas e estruturais de folhas e perfilhos foi um método interessante para agrupar cultivares, demonstrando a importância e o potencial de uso da morfogênese em trabalhos de seleção e avaliação de plantas forrageiras.
Palavras-chave:Brachiaria, Panicum, variáveis morfogênicas e estruturais
Introduction
During the last decade there were significant changes and advances in the comprehension of facts and processes related to the use of forage grasses in tropical grasslands (Da Silva and Nascimento Júnior, 2007; Nascimento Júnior et al., 2002). Several of these changes originated from morphogenesis studies carried out with a variety of forage grasses in different geographical locations and using different grazing methods, whose objective was to understand aspects relative to the form and function of plants responsible for their grazing management requirements. This kind of information is still not abundant given the difficulties related to conducting grazing studies and the large, increasing number of forage plant options available to farmers.
In spite being laborious, morphogenesis studies allow tracking of the turnover dynamics of leaves and tillers, basic components of the herbage accumulation process (Nascimento Júnior, et al., 2004). They also provide an insight of the general pattern of plant growth, contributing to a better understanding of plant functionality and generating basic knowledge necessary to plan and define adequate grazing management strategies (Barbosa et al., 2002). Additionally, studies of this nature make possible the identification of plants with contrasting production potentials, since some variables like leaf and stem elongation rate are highly correlated with herbage mass and accumulation rates as well as allow grouping of plants with similar patterns of growth (functional groups) and management requirements regardless of plant species or cultivar.
The objective of this experiment was to carry out a comparative study of ten tropical forage grasses based on plant morphogenetic and structural responses aiming at identifying groups of cultivars with similar functionality and, therefore, similar management requirements. This would allow a generalisation of management principles across a range of plant types, allowing inferences to be made about management of plants with similar patterns of growth from other species and/or cultivars.
Material and Methods
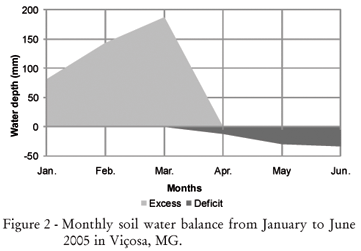
The experiment was carried out in Viçosa, state of Minas Gerais, Brazil (20º45'S; 42º51'W; and 651 m altitude). According to the classification of Köppen, the weather type is Cwa, subtropical, with a well-defined dry season during the winter months and a rainy season during summer. The monthly average maximum, medium and minimum air temperatures as well as rainfall are shown in Figure 1, and a monthly extract of the soil water balance (Thornthwaite and Mather, 1955) during the experiment is presented in Figure 2.
Local soil is classified as Inceptic Hapludults, with clay-loam texture. Results from chemical analysis of soil samples (0-20 cm) revealed a pH of 5.82, no aluminium saturation, base saturation of 41.2%, phosphorus and potassium concentrations (Mehlich-1) of 1.79 and 24.0 mg dm3. At planting, only phosphorus fertiliser was used at a rate of 110 kg ha1 of P2O5, applied as simple superphosphate on planting rows. Nitrogen and potassium fertiliser were applied as urea and potassium chloride after the complete establishment of plants, at rates of 50 kg ha1 of N and 90 kg ha1 of K2O, divided in two broadcast applications.
The experiment started on Dec 12, 2004 and finished in Jun 30, 2005, totaling 178 days of experimental period. Treatments corresponded to ten grass cultivars of which two were from the Panicum genus (P. maximum cultivars Tanzânia and Mombaça) and eight from the Brachiaria genus (B. Brizantha cultivars Piatã, Marandu, Xaraés, Capiporã and Arapoty; B. decumbens cultivar Basilisk; B. humidicola cultivars Comum and Tupi). Cultivars seeds were sown in plastic tubes containing commercial organic substrate in glasshouse conditions. Plants were then transplanted from the tubes to field plots according to a 0.15 m distance between plants in each row and 0.20 m distance between rows. The experimental design consisted of a randomised complete block, with three replicates, totaling thirty 0.90 m2 plots (0.90 m × 1.00 m).
Because cultivars had contrasting growth forms (prostrated, semi-prostrated and tall-tufted grasses) and consequently defoliation management requirements, plots were staged prior to the commencement of the experiment and plants were monitored under free growth conditions. Evaluations were performed during the entire period of the vegetative development of the plants and were interrupted when 50% or more of the plants in the plot had visible flower heads.
Measurements of morphogenetic and structural variables were performed twice a week on two tillers randomly chosen and tagged per tussock in each plot. As soon as marked tillers produced flower heads, new vegetative tillers were tagged, but monitoring of reproductive tillers was not interrupted. Data were used to calculate the following variables: leaf appearance rate (LAR leaves tiller1 day1), phyllochron (Phyl days leaf1 tiller1), leaf elongation rate (LER cm tiller1 day1), stem elongation rate (SER cm tiller1 day1), final leaf length (FLL cm tiller1), number of live leaves per tiller (NLL leaves tiller1) and leaf life span (LLS days).
Measurements related to tiller demography and tillering dynamics were performed on two randomly chosen tussocks per plot, according to the methodology described by Carvalho et al. (2000). Tillers on the chosen tussocks were tagged with plastic coated wires of different colours every 30 days and sorted out into basal and aerial tillers. At the end of the experiment seven generations of tillers had been monitored. The dead tillers, by generation, were also accounted for, allowing calculation of the rates of tiller appearance, death and survival.
Data related to descriptive variables such as tiller demography patterns, number of new leaves produced (NNL), number of live leaves (NLL) and number of dead leaves per tiller (NDL) are presented using descriptive statistics only. Analysis of variance was performed on morphogenetic and structural variables using the GLM procedure of SAS (SAS Institute, 1996), considering a split-plot arrangement where grass cultivars were allocated to plots and time to subplots. The commands RANDOM, TEST and LSMEANS were used to identify and to carry out appropriate tests. Comparison between treatment means, when necessary, was made using Tukey test and a 5% significance level.
In order to identify common features and patterns of growth among cultivars, a grouping analysis was performed according to the methodology proposed by Tocher, cited by Rao (1952) and described in Cruz and Regazzi (1997) using the morphogenetic and structural variables of leaves and tillers in an attempt to identify functional groups of plants with similar patterns of plant development and management requirements regardless of plant species and/or cultivar.
Results and Discussion
The results for morphogenetic and structural variables (Table 1) show that leaf appearance rate varied with grass cultivars (p = 0.0009), with the highest value recorded for Tupi (0.22 leaf tiller1 day1). The lowest value was recorded for Capiporã (0.08 leaf tiller1 day1), and there was no difference relative to values recorded for cultivars Arapoty, Marandu, Piatã, Basilisk, Xaraés, Mombaça and Tanzânia.
Phyllochron results followed the same pattern of LAR results (p = 0.0182), since they were calculated as the inverse of LAR. Higher values were recorded for cultivars Capiporã, Xaraés and Piatã (13.68, 12.88 and 12.72 days leaf1 tiller1, respectively) relative to the remaining cultivars. As expected, the cultivar Tupi showed the lowest Phyl (5.50 days leaf1 tiller1), not differing from the cultivars Comum and Mombaça.
LER varied with grass cultivars (p < 0.0001), with highest values recorded for cultivars Mombaça and Tanzânia (5.79 and 5.23 cm tiller1 day1, respectively) and lowest values for cultivars Comum, Basilisk and Tupi (1.29, 1.72, 2.05 and cm tiller1 day1, respectively). This is an important variable in studies regarding morphogenetic responses and dynamics of herbage accumulation, since it has a high positive correlation with sward herbage mass (Horst et al., 1978). As a result, the two cultivars of B. humidicola (Comum and Tupi) and of B. decumbens (Basilisk) presented the lowest potential of leaf accumulation, the opposite of the two cultivars of P. maximum (Mombaça and Tanzânia). When cultivars had low potential of leaf accumulation because of their low rate of leaf elongation, they usually had higher rates of leaf appearance, suggesting a smaller period of leaf elongation and a higher turnover of leaves, as was the case for Tupi.
Cultivars also differed in terms of FLL (p < 0.0001), with highest values recorded for cultivars Tanzânia (54.44 cm tiller1) and Mombaça (52.26 cm tiller1) of P. maximum. On the other hand, the lowest values of FLL were recorded for cultivars Tupi, Comum and Basilisk (11.65, 12.39 and 14.41 cm tiller1, respectively). According to Nabinger and Pontes (2001), while plants with high LER usually present long leaves, plants with short leaves are usually characterised by high LAR. In this experiment, the cultivar Tupi had the highest LAR, and because of the inverse relationship between LAR and LER, it was among the three cultivars with the lowest LER. The association between these two variables determines FLL and, in fact, cultivars with higher LAR and, consequently, lower LER, presented lower FLL.
SER varied with grass cultivars (p = 0.0124), with the highest value recorded for the cultivar Tupi (2.01 cm tiller1 day1) and the lowest for Capiporã (0.33 cm tiller1 day1). SER for cultivar Tupi was calculated based on monitoring of stolons. Cultivars also differed (p = 0.0061) in terms of LLS, with higher values recorded for cultivars Piatã (78 days), Xaraés (74 days), Mombaça (74 days), Tanzânia (72 days) and Capiporã (63 days) and lower for Tupi (43 days), Arapoty (45 days) and Comum (47 days) . These results indicate a higher leaf turnover for cultivars Tupi, Comum and Arapoty of Brachiaria, since they had also high values of LAR. Since the number of live leaves per tiller is a genetic characteristic, when plants are managed within agronomic limits of plant tolerance and resistance to defoliation, values are relatively stable for each plant species and/or cultivar (Davies, 1974). In this case, it would be only natural to expect that plants with higher LAR would have to have lower LLS.
NLL varied with grass cultivars (p = 0.0107), with highest value recorded for the cultivar Tupi (8.0 leaves tiller1), probably a result of its high LAR. Cultivar Basilisk, on the other hand, had the lowest NLL (3.4 leaves tiller1). The number of leaves per tiller of plants from the genus Panicum and Brachiaria throughout the experiment shows that, in general, cultivars of B. brizantha presented more similarities than differences in relation to cultivars of P. maximum, while cultivars of B. humidicola have more similarities than differences in relation to the cultivar of B. decumbens.(Figure 3). NLL for cultivars of P. maximum, B. brizantha, B. decumbens and B. humidicola decreased as the experiment progressed and this was associated with a progressive increase in leaf death. Cultivars of P. maximum had an average of 6.4 live leaves per tiller, while cultivars B. brizantha, B. decumbens and B. humidicola showed 5.0, 3.3 and 6.0 live leaves per tiller, respectively. NLL is a characteristic that remains relatively constant throughout the plant cycle within a range of environmental and management conditions (Lemaire and Chapman, 1996). In this study the decrease in NLL as the experiment progressed was probably associated to the beginning of the reproductive development of plants, a condition characterised by the cessation of leaf appearance and increase in leaf mortality.
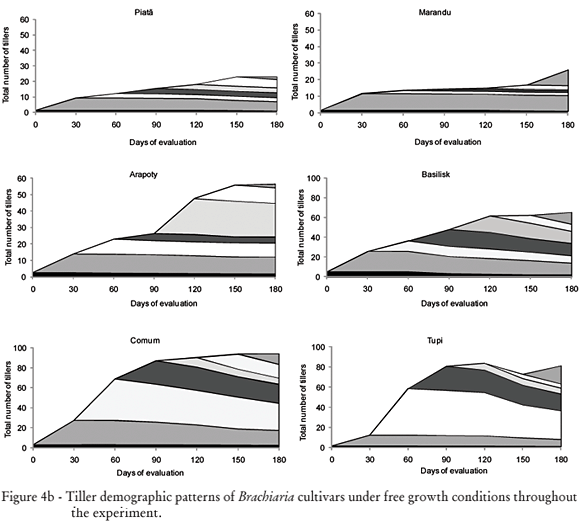
In relation to patterns of tiller demography (variation in tiller numbers throughout time as a function of tiller appearance and death)(Figures 4a and 4b), cultivars Tanzânia, Mombaça and Xaraés presented similar patterns, with tiller population being comprised mainly by basal tillers. Cultivar Capiporã did not produce aerial tillers, while the number of tillers of cultivar Capiporã was similar to that of cultivars Tanzânia, Mombaça and Xaraés. This indicates that differently from other cultivars, tiller population of the cultivar Capiporã was mainly formed by tillers from generations 1, 2, 3 and 4, with little or none participation of tillers from generations 5, 6 and 7.
For cultivars Piatã and Marandu there was a substantial contribution of aerial tillers to tiller population. Cultivar Arapoty had a very distinct pattern of tiller demography in relation to the other cultivars of B. brizantha, characterized by a higher tiller population and also a higher contribution of aerial tillers. Cultivar Basilisk presented a pattern of tiller demography similar to those of the cultivars Comum and Tupi. It was for these three cultivars that the highest values of tiller population were recorded, all with a significant contribution of aerial tillers. It should be pointed out that the appearance of basal and aerial tillers occurred simultaneously, in those mentioned cultivars for all generations.
In general, the appearance of new tillers was high at the beginning of the experiment, probably a consequence of the sward still being established. The second and/or the third generation of tillers were those that most contributed to final tiller population for the cultivars Mombaça, Tanzânia, Xaraés and Capiporã. The significant contribution of these generations can also be attributed to favorable factors at the time of establishment as, for example, high temperatures and rainfall (Figures 1 and 2).
Analysis of the data using the Tocher method allowed the identification of five groups of grass cultivars (Table 2), suggesting a potentially important application of morphogenesis in studies of this nature. The cultivar Basilisk of B. decumbens was placed into the same group as cultivar Comum of B. humidicola, the same happening for cultivar Xaraés of B. brizantha and cultivars Mombaça and Tanzânia of P. maximum, indicating that in spite of belonging to different plant species, Comum and Basilisk as well as Xaraés, Mombaça and Tanzânia had similar growth patterns and/or functionalities. The Mahalanobis generalised distance within and between groups are shown in Table 3.
The Mahalanobis generalised distance between groups 1 and 3 was 14.9068. The distance between groups 4 and 5 was small (D2 = 4.3561), indicating that in spite of being two separate groups, they had similar patterns of growth, as described by the variables in Table 1 and verified by the similarity presented by the plants of this group in relation to the tillering demography (Figures 4a and 4b). The higher distance between groups 1 and 3 in comparation to groups 4 and 5 can be explained by the fact that the cultivars of groups 1 and 3 belong to different genders, but, in spite of this, the cultivars presented similar functionality in relation, for instance, to leaf appearance that was low for these two groups, leaf elongation was high (Table 1), as well as, NDL (Figure 3) that was high for the plants of that group.
Some of the groups formed were comprised by plants from different species, indicating that despite differences in morphology plants within groups had similar patterns of growth and development. This opens a new window of opportunities in terms of research and management of tropical grasses, which would make it possible to define general principles of management in terms of groups of plants with similar functionality and allow inferences to be made about management requirements of new potentially interesting plants taking into account their patterns of growth. As an example, the group comprised by Tanzânia, Mombaça and Xaraés (group 1) was characterised by plants with high potential of herbage accumulation (low values of LAR and high to medium values of LER), tussock plants with smaller number of heavier tillers. Plants in this group have long leaf life span and, for that reason, suitable for management using longer defoliation intervals relative to the groups comprised by Tupi, Basilisk and Comum (groups 4 and 5), characterised by plants with lower potential for herbage accumulation (high values of LAR and low values of LER), with great number of small tillers, lower leaf life span and, therefore, suitable for management using shorter defoliation intervals.
Conclusions
In spite of the differences among cultivars considering all variables and the variability of the plant material, plants can be grouped in terms of similar growth patterns. This would be done regardless of plant species and/or cultivar, allowing general principles of defoliation management to be developed and inferences to be made about management requirements and potential for herbage accumulation of new potentially interesting plant material, indicating the importance that morphogenetic traits may have in forage plant breeding and selection programmes.
Acknowledgements
To FAPEMIG and to the Animal Science Department of the Federal University of Viçosa for the opportunity and sponsorship of the first author. Thanks are also due to the teammates: Anderson de Moura Zanine, Bráulio Maia de Souza, Karine da Silva Pena and Wilton Ladeira da Silva, for their help in carrying out the research project, and to Antônio Augusto Bianchi for the English version of this article.
Received October 02, 2008
Accepted December 22, 2009
- Barbosa, R.A.; Nascimento Júnior, D.; Euclides, V.P.B.; Regazzi, A.J.; Fonseca, D.M. 2002. Morphogenic characteristics and forage accumulation of Tanzania grass (Panicum maximum Jacq. cv. Tanzânia) in two pos grazing residues. Revista Brasileira de Zootecnia 31: 583-593. (in Portuguese, with abstract in English).
- Carvalho, C.A.B.; Da Silva, S.C.; Carnevalli, R.A.; Sbrissia, A.F.; Pinto, L.M.; Fagundes, J.L.; Pedreira, C.G.S. 2000. Tillering and forage accumulation of a Florakirk pasture (Cynodon spp.) managed in four canopy heights. Boletim da Indústria Animal 57: 39-51. (in Portuguese).
- Cruz, C.D.; Regazzi, A.J. 1997. Biometrics models applied to genetic breeding. UFV, Viçosa, MG, Brazil. (in Portuguese, with abstract in English).
- Da Silva, S.C.; Nascimento Júnior, D. 2007. An overview of the research on tropical grasses: morphogenic characteristics and grazing management. Revista Brasileira de Zootecnia 36: 122-138. (in Portuguese, with abstract in English).
- Davies, A. 1974. Leaf tissue remaining after cutting and regrowth in perennial ryegrass. Journal of Agricultural Science 82: 165-172.
- Horst, G.L.; Nelson, C.J.; Asay, K.H. 1978. Relationship of leaf elongation to forage yield of tall fescue genotypes. Crop Science 18: 715-719.
- Lemaire, G.; Chapman, D. 1996. Tissue flows in grazed plant communities. p. 3-36. In: Hodgson, J.; Illius, A.W., eds. The Ecology and Management of Grazing Systems. CAB International, Wallinford, UK.
- Nabinger, C.; Pontes, L.S. 2001. Morphogenesis of forage plants and sward structure. p. 755-771. In: Mattos, W.R.S., ed. Proceedings of the Annual Meeting of the Brazilian Society of Animal Sciences 38. SBZ, Piracicaba, SP, Brazil. (in Portuguese).
- Nascimento Júnior, D.; Da Silva, S.C.; Adese, B. 2004. Future perspectives of grasses use under grazing. p.130-141. In: Medeiros, S.R.; Euclides Filho, K.; Euclides, V.P.B., eds. Proceedings of the Annual Meeting of the Brazilian Society of Animal Sciences 31., SBZ, Campo Grande, MS, Brazil. (in Portuguese).
- Nascimento Júnior, D.; Neto, A.F.G.; Barbosa, R.A.; Andrade, C.M.S. 2002. Bases for the grazing management: an overview. p. 149-196. In: Obeid, J.A.; Pereira, O.G.; Fonseca, D.M.; Nascimento Júnior, D., eds. Proceedings of the Symposium on Strategic Management of Pasture 4. SIMFOR, Viçosa, MG, Brazil. (in Portuguese).
- Rao, R.C. 1952. Advanced statistical methods in biometric research. Wiley, New York, NY, USA.
- SAS Institute. SAS/STAT 1996: user's guide statistics. 6.4ed. SAS Institute, Cary, NC, USA.
- Thornthwaite, C.W.; Mather, R.J. 1955. The water balance. Drexel Institute of Technology, Centerton, NJ, USA.
Publication Dates
-
Publication in this collection
26 Apr 2010 -
Date of issue
Apr 2010
History
-
Received
02 Oct 2008 -
Accepted
22 Dec 2009