Abstracts
Knowledge acquired about the main immune mechanisms protecting against various infectious agents leads to a better understanding of the pathogenesis of infectious/parasitic diseases and of various strategies specific to the host and parasite. The immune system performs along a cooperation network, which involves the participation of several structural, molecular and cellular components. The fine balance between health and disease is found in this scenario, in which deficiency as much as excess may result in tissue damage. This article explores such aspects and a number of therapies arising from the knowledge acquired.
helminthiasis; immunity; immunity; infection; bacterial infections; protozoan infections; mycoses; virus diseases
O conhecimento dos principais mecanismos de defesa imune contra os diversos agentes infecciosos permite a compreensão da patogênese das doenças infectoparasitárias e das várias estratégias do hospedeiro e do parasita. O sistema imunológico atua numa rede de cooperação, envolvendo a participação de muitos componentes estruturais, moleculares e celulares. Nesse cenário encontra-se o delicado equilíbrio entre a saúde e a doença, em que tanto a deficiência quanto o exagero resultam em dano tecidual. Este artigo explora esses aspectos e algumas abordagens terapêuticas que surgem desse entendimento.
helmintíase; imunidade; imunidade natural; infecção; infecções bacterianas; infecções por protozoários; micoses; viroses
CONTINUING MEDICAL EDUCATION
Immune response mechanisms to infections* * Work done at the Immunology Service, Prof. Edgard Santos University Hospital. Federal University of Bahia (UFBA), Salvador, Bahia, Brazil.
Paulo R. L. MachadoI; Maria Ilma A. S. AraújoII; Lucas CarvalhoIII; Edgar M. CarvalhoIV
IPh.D. in Medicine. Research fellow of the UFBA Immunology Service. Assistant Professor of Dermatology, Baianese Foundation for Development of the Sciences
IIPh. D. in Cellular and Molecular Biology. Research fellow of the UFBA Immunology Service. Assistant Professor of Dermatology, Baianese Foundation for Development of the Sciences
IIIPh.D. in Patology. Research fellow of the UFBA Immunology Service
IVTitular Professor of Medicine, UFBA. Head of the Immunology Service, UFBA
Correspondence Correspondence to Paulo R. L. Machado Serviço de Imunologia - 5º andar Hospital Universitário Prof. Edgard Santos UFBA Rua João das Botas s/n - Canela Salvador Bahia 40110-160 Fax: (71) 245-7110 E-mail: prlmachado@uol.com.br
ABSTRACT
Knowledge acquired about the main immune mechanisms protecting against various infectious agents leads to a better understanding of the pathogenesis of infectious/parasitic diseases and of various strategies specific to the host and parasite. The immune system performs along a cooperation network, which involves the participation of several structural, molecular and cellular components. The fine balance between health and disease is found in this scenario, in which deficiency as much as excess may result in tissue damage. This article explores such aspects and a number of therapies arising from the knowledge acquired.
Key-words: helminthiasis; immunity; immunity, natural; infection/immunology; bacterial infections; protozoan infections; mycoses; virus diseases.
INTRODUCTION
Immune response plays a vital role in protecting against infectious agents. It is the main impediment against the occurrence of disseminated infections that are usually associated with a high death rate. It is a well-known fact that for virtually all infectious diseases, the number of individuals exposed to infection is much higher than those actually presenting with a disease. This indicates that most persons are able to destroy these microorganisms and thus prevent the progression of an infection. By contrast immune deficiencies, whether of innate immunity (phagocytic cell dysfunction or complement deficiency) or adaptive immunity (antibody production deficiency or T-cell function deficiency), are strongly associated with increased susceptibility to infections.1
Although immune response is fundamental for protecting against most infectious agents, evidence has been accumulating over the years as to how in many infectious diseases the main pathological aspects are not related to the direct action of an aggressor agent, but instead to abnormal immune response. In several such situations, there exists a hypersensitivity reaction with exaggerated and non-modulated immune response, the result of which is tissue damage. In other cases, infectious agents, whether by mimicking the antigens themselves, by inducing a proliferation of self-reactive cells or by increasing the expression of MHC and co-stimulatory molecules in infected cells can precipitate autoimmune diseases.2
That different types of microbes are combated by different immune response components has been known since the beginning of the 1950s, when the importance of antibodies on the destruction of extracellular bacteria was first documented. Although antibodies on their own and in isolation are not able to destroy bacteria, they may neutralize microorganisms by preventing them from binding to the host tissue. Furthermore, in association with the complement, antibodies may lyse bacteria and function as opsonins, thereby facilitating phagocytosis. Neutrophils, eosinophils and macrophages exert their microbicid action most broadly against various types of agents and are extremely important cells for protecting the host. Documentation on how phagocytic cells express their receptor membranes like the toll-like receptor (TLR), which binds specifically with existing molecular patterns in various infectious agents,3 makes it inaccurate to name innate immune response non-specific. Neutrophils play a fundamental microbicid role against bacteria; macrophages are important cells for defending against intracellular agents (protozoan and intracellular bacteria). Eosinophils are essential not so much due to phagocytic activity as to cytotoxic activity against helminthes. T-cell mediated response is highly effective for the protection mechanism against intracellular agents, like viruses, protozoans, funguses and intracellular bacteria. T-cells may exercise their function through cytotoxicity mediated by CD8+ cells or through the secretion of cytokines, which activate macrophages to destroy intracellular agents. Other elements that may participate in the process of protecting against infectious agents included keratinocyte and Langerhans cell, since the skin is often invaded by various microorganisms. Keratinocytes are able to secrete innumerable cytokines, thereby activating and recruiting inflammatory cells and lymphocytes for the skin.4 The Langerhans cell in turn exercises a fundamental role of watching over the cutaneous territory, and phagocyting everything from particular inanimate proteins up to and including viruses, bacteria or other microorganism invaders. After phagocytosis the Langerhans cell migrates to the regional lymph node to carry out the antigenic presentation of the lymphocytes, which begins the development of specific protector immunity, tolerance or hypersensitivity.5
The cells and mediators involved in protecting humans are well known. Yet the fact that TCD4+ (T helper) are heterogeneous and made up of two subpopulations, namely Th1 and Th2 cells, has only recently been documented.6 This observation has contributed a lot to understanding the immunopathogenesis of the most infectious diseases. Figure 1 shows the dichotomy of the TCD4+ cells and mediators produced by them.
It is vital to understand that both Th1 and Th2 responses are important in the task of protecting the host against infection. Th1 response is related to protecting against protozoans, intracellular bacteria and viruses, while Th2 response is more effective against helminthes and extracellular bacteria. These responses are also antagonistic, insofar as the IFN-gamma negatively modulates Th2 response, and IL-4 and IL-10 negatively modulate Th1 response. This enables homeostasis in the immune system and a balanced immune response. In addition, the regulatory cells of immune response which express molecules CD4 and CD25 (Tr) and produce IL-10 and/or TGF-beta (Tr1 or Th3) are involved in modulating immune response. They prevent or reduce the consequences of hypersensitivity reactions and auto-immune diseases.7
1. IMMUNE RESPONSE AGAINST BACTERIA
Bacteria are the microorganisms that most frequently cause infections in humans. The natural barriers against infection agents as well as innate and adaptive immunity participate in the protection mechanism against bacteria.
1.1. Intracellular Bacteria
The main characteristic is the ability to survive within the macrophages, for example M. tuberculosis, M. leprae and L. monocitogenesis. Penetration into the macrophage also constitutes the parasite's escape mechanism. Although paradoxical, the latter is benign for the host insofar as the lack of cellular penetration by the bacteria may induce a strong inflammatory response and excessive damage for the host. Within the macrophages these bacteria may stimulate either TCD4+ cells by an expression of the antigen associated to MHC class II or TCD8+ cells by an expression of the antigens associated with molecules of MHC class I. Activation of TCD4+ cells lead to the secretion of IFN-gamma, which activates the macrophages and leads to increased production of nitrous oxide (NO) and destruction of bacteria. TCD8+ cells participate in the protection mechanism through cytotoxicity, thereby destroying the infected macrophages. In the case of M. tuberculosis, despite having immune protection preventing its multiplication, there is no complete elimination of the bacillus. For this reason, individuals using corticosteroids and HIV-positive patients develop clinical signs of tuberculosis, despite having been infected much earlier and after remaining completely asymptomatic. The role of cellular immune response in controlling infections caused by mycobacteria is well demonstrated in how these infections have spread with the advent of AIDS.
Regarding infections caused by M. leprae, the clinical spectrum of the disease is intimately linked to immune response. In patients having a tuberculoid form, there is a strong response to Th1. Also, the disease is characterized by destruction of the nervous fibers in specific areas leading to the appearance of localized and well-delimited skin lesions, with a loss of sensitivity to heat and pain. When Th1 response is lacking, there is a dissemination of of the bacillus, which leads to Virchowian Hanseniasis. In this event, the macrophages are replete with the parasite and there is a thickness of lymphocytes found on the lesion. Borderline forms, also known as dimorphic, represent a clinical and immunological pattern of intermediary response. 8
The importance of immune response in Hanseniasis disease is not limited to the determination of its clinical spectrum. With the onset of disease or often after treatment is started, some patients may show acute secondary clinical signs after the release of antigens and hypersensitivity reactions. These manifestationsalso called reactionsare represented by erythema nodosum leprosum (ENL) and reverse reaction (RR). ENL is a systemic inflammatory response associated with high concentrations of tumor necrosis factor alpha (TNF-alpha) and the deposition of immunocomplexes with an infiltration of neutrophils and the activation of a complement, involving various organs.9,10 The immunopathogenesis of ENL is quite complex. High levels of circulating IL-1 and TNF-alpha,11,12 have been found in patients' feces, whereas a tissue increase in the expression of messenger RNA by IL-6, IL-8 and IL-10 indicates Th2 response.10,13 Moreover, the presence of the inductible nitrous oxide synthase enzyme (iNOS) has been documented as potentially being induced in the neutrophils and TNF-alpha and TGF-beta in the macrophages of the lesions.14 ENL may be accompanied by systemic toxicity, which is often treated with corticosteroids or TNF-alpha inhibitory drugs, like thalidomide. On the other hand, RR develops in the wake of the abrupt emergence of a delayed hypersensitivity mechanism against antigenic fractions of M. leprae, involving the active participation of T lymphocytes with tissue production of Th1 cytokines (IL-2, IFN-gamma) and inflammatory cytokines, like TNF-alpha.13 The lesions appear to be infiltrated by CD4+ lymphocytes, with increased expression of HLA-DR and of the receptor IL-2 in cells of the infiltrate, just as with those in the keratinocytes.15
1.2. Extracellular Bacteria
Infections caused by extracellular bacteria are the most frequent of all. In these cases, the protection mechanisms are mainly related to the host's natural barriers, innate immune response and antibody production.
The importance of natural barriers in the fight against extracellular bacterial infections is well known. The integrity of skin and mucosas prevent adherence and penetration of bacteria; mucociliar movement eliminates bacteria from the respiratory tract; the stomach's acidic pH destroys bacteria penetrating by the upper digestive tract; and in the saliva and prostatic secretions there exist substances with antimicrobial activity. Chart 1 provides details of the main protection mechanisms against extracellular bacteria.
The participation of innate immunity occurs through phagocyte cells, the activation of a complement system through an alternative path and by production of chemokines and cytokines. In addition, C-reactive protein (CRP), an acute phase protein produced mainly by hepatic cells in bacterial infections, exerts a diversified range of action against the bacteria. When binding to phospholipids of the membrane of some bacteria (for example, pneumoccocus) CRP works like opsonin, facilitating the phagocytosis by neutrophils. CRP also has the capacity to activate the complementary system and stimulates the synthesis of TNF-alpha, which induces the synthesis of NO and consequently the destruction of various microorganisms.
The complement performs its protection role by activating the attack complex at membrane (C5-C9) and facilitates opsonization through the C3b component, which binds to the bacteria and interacts at a second stage with the specific receptor existing in phagocytic cells. The deficiencies of the complementary system have been associated with serious infections by Neisseria meningitides and infections disseminated by Neisseria gonorheae.16
All innate immunity cells participate in protecting against bacteria, though it is the role of neutrophils and monocytes/macrophages that are mainly emphasized by the phagocytic capacity of these cells. The basophiles and mastocytes activated by factors of the complement system, as in C5a, C3a and C4a for example, release mediators which, when combined with the aforementioned complement proteins, attract leukocytes to the site of aggression and contribute to the passage of these cells from the vessels to the tissues, namely the site at which the aggression against the host occurs. Apart from its phagocytic activity, eosinophils may destroy microorganisms by means of releasing proteins with microbicid activity, such as the main basic protein and eosinophil cationic protein. Neutrophils and macrophages play a key role in protecting against these agents provided that bacteria are susceptible to substances produced by these cells, for example NO and hydrogen peroxide. Within these cells, enzymes like myeloperoxidase and other substances like azurocidin having microbicid properties also exist. Although neutrophils as well as macrophages are phagocytic cells, they have much different characteristics. Whereas neutrophils have a short lifespan in either the blood or tissues, macrophages survive over extended periods of time. Neutrophils are only found in inflamed tissues, while macrophages are concentrated either in inflamed or healthy tissues. During the inflammatory reaction, neutrophils produce purulent secretion, whereas the macrophages form granuloma. Neutrophils mainly protect against extracellular bacteria, whereas macrophages are vital to eliminate the intracellular agents that house them.
Immune response cells are also the main sources of cytokines and chemokines at the onset of the infection. They exert inhibitory action either on the innate or adaptive phase. Due to their role of attracting cells to the lesion site, chemokines are very important in the process of protecting the host.17
Among the various cytokines that participate in protecting against bacteria, the pro-inflammatory cytokines, like TNF-alpha, IL-1 and IL-6, are noteworthy. These cytokines are produced in the initial phases of the infection. By means of their action on the hypothalamus, they are responsible for the appearance of a fever that inhibits bacterial multiplication. They increase the expression of adhesion molecules (Seletine P and ICAM), thereby easing the passage of cells from the vessel to the infection site. They also stimulate neutrophils and macrophages to produce NO and destroy bacteria. Other cytokines produced in the initial infection phases interfere with the adaptive immune response. Produced by macrophages, IL-12 has an important role in the differentiation of Th0 cells into Th1 cells.18 By contrast, IL-4, produced by basophiles, mastocytes and macrophages, stimulates a differentiation of Th0 cells into Th2 cells, which end up collaborating with lymphocyte B in the production of antibodies, but especially of IgE.19
Adaptive immunity, mainly by means of antibodies, performs an important role against these extracellular bacteria. The antibodies may perform their inhibitory action in three ways: 1) opsonization, 2) activating the complement system, 3) promoting the neutralization of bacteria or its products.
Extracellular bacteria are susceptible to destruction when phagocyted. They develop substances like the evasive mechanism that have antiphagocytic activity. Antibodies aimed against these substances not only impede upon their action, but facilitate phagocytosis, insofar as the neutrophils and macrophages have receptors for the FC portion of the immunoglobulin (opsonization). Antibodies also co-assist in destroying bacteria by the complement, and activate this system by a classic pathway. By means of the neutralization mechanism, the antibodies, primarily IgA, may bind with the bacteria and accordingly prevent the latter from establishing themselves in the mucosas, intestinal tract and respiratory tract. Antibodies often bind to bacteria-produced toxins, like tetanic and diphtheric toxins, and neutralize the action of these products.
Despite the protective importance of immune response, the difficulty in controlling the inflammatory response that develops may provoke tissue damage, which is nonetheless most often limited and without greater consequences for the host. However, infections caused by gram-negative germs may eventually result in septicemia and septic shockvery serious situations usually associated with a high mortality rate. Septic shock is triggered by lipopolyssacharides (LPS) present in the bacterial wall, which stimulate an exacerbated production of pro-inflammatory cytokines in the neutrophils, macrophages, endothelial cells and muscles (TNF-alpha, IL-1, IL-6, IL-8) and NO. Muscle tone and heart beat are reduced as a result, which leads to hypotension and poor tissue perfusion, and finally cellular death. By contrast, modulation of this exacerbated response may be obtained. As such, in an experimental model, the concomitant combination of IL-10 and LPS protects mice from death during septic shock by inhibiting the production of IL-12 and synthesis of IFN-gamma and TNF-alpha.20
2. IMMUNE RESPONSE IN VIRAL INFECTIONS
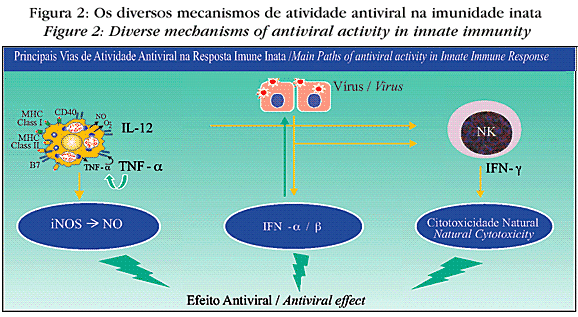
Despite the manifold mechanisms of protecting against viruses, viral diseases are not only common, but in fact represent one of the most important infectious diseases today associated with mortality in the general population. Figure 2 shows how viruses are destroyed by means of innate immune response. In the initial phase of viral infections, controlling the infections is done with interferons type I (IFN-alpha and IFN-beta), macrophages and NK cells.21
Type I interferons are produced by virus-infected cells. By interacting with a non infected cell, their feature is to protect them against infection in addition to collaborating with adaptive immune response. IFN-gamma also acts against virus infections by means of activating the macrophages to destroy the virus as well as the NK cells (natural cytotoxic cells) to release granzyme and perforin and destroy infected cells. In addition, IL-12 plays an important part in the initial phase. It is produced by macrophages and other antigen-presenting cells. It stimulates NK to exert cytotoxicity and produce more IFN-gamma, which in turn increases the microbicid potential of macrophages.
Adaptive immunity against viral antigens occurs with the activation of TCD8+ cells that exert cytotoxicity when recognizing viral antigens via MHC class I in the target cells, with a result of releasing granzyme and perforins with the lysing of the infected cells and virus. During adaptive immune response TCD4+ cells are also activated, which then go on to collaborate with B cells to produce antibodies. In spite of viruses being intracellular agents, antibodies play an important role in fighting against viral infections insofar as the viruses break open these cells and remain free until penetrating into another cell. In this extracellular phase, antibodies may bind to the virus, and by means of the neutralization of the mechanism, prevent others from penetrating a non infected cell. By contrast, antibodies may assist in the cellular cytotoxicity mechanism that depends on them, by binding to the infected cells and thereby allowing NK cell action. In various diseases, as in the examples of poliomyelitis, measles, hepatitis B and varicella, the antibody has a fundamental role in protecting against infection when it is a previously sensitized host, whether by a prior infection or immunization. This is because, in already sensitized individuals, the presence of antibodies can intercept the virus and thus prevent it from binding to the host cell.
In virtue of several protective mechanisms against viruses, a large part of viral infections are asymptomatic or have a subclinical presentation with non specific manifestations, like fever and cutaneous rash. Nonetheless, various viral infections do progress and important tissue damage can occur. The pathology associated with viral infection may be related to the virus' cytopathic effect, hypersensitivity reaction and auto-immune phenomena (Figure 3).
In many viral infections, cells are destroyed through a process involving more than one of these mechanisms. For example, in HIV-infection and infections by hepatitis viruses B and C, the destruction of infected cells is mediated as much by the virus' cytopathic effect as through cytotoxicity by NK and CD8 cells. Some viral infections perfectly exemplify the broad dimension of aggression mechanisms occurring against tissue in the course of these infections.
2.1. Human Immunodeficiency Virus (HIV)
HIV infects TCD4+ cells predominantly. The destruction of these cells may occur by the virus' cytopathic effect. In addition, there exists increased apoptosis in these cells. Due to expressing viral antigens at the level of the membrane, the cells may also be destroyed by cytotoxicity mediated by the TCD8+ cell, a phenomenon also contributing to the reduction of CD4+ cells. As the CD4+ cell is one of the most important for obtaining the cooperation of immune response, the numerical reduction and alteration of its function leads to the suppression of immune response. This suppression is associated predominantly with a reduction of IL-2, IFN-gamma and TNF-alpha.22 This is why in AIDS patients, the main opportunistic infections are related to intracellular agents such as: M. tuberculosis, P. carinii, cytomegalovirus, C. albicans and criptosporidium. As in HIV infection, memory B lymphocytes keep functioning, antibodies are produced and the protection mechanism against extracellular agents does not experience large scale damage. However, this lack of greater susceptibility to extracellular bacterial infections observed in AIDS patients is observed in adults in whom the repertory of B-cell produced antibodies depending on T-cells had already formed prior to HIV infection. In infected children, as the alteration of TCD4+ cell functioning is premature, cellular cooperation is damaged with abnormalities also occurring in the synthesis of antibodies. This is why infections by extracellular bacteria are common in HIV-infected children.
2.2. Human T cell Lymphocytotropic virus (HTLV-1)
Infection by the HTLV-1 induces activation and intense cellular proliferation of infected T lymphocytes. This phenomenon is related mainly to the function of the virus' Tax gene, whose property is to transactivate IL-2 and IL-2-receptor genes. These T-cell proliferation anomalies may lead to the appearance of leukemia in adult T cells. Indiscriminate cell proliferation may also provoke an expansion of self-reactive T cells and accentuated secretion of pro-inflammatory cytokines like TNF-alpha. These abnormalities may associate with cutaneous and neurological tissue lesions.23
Owing to the strong Th1 cell activation in HTLV-1 infection, there is reduced production of IL-4 and IL-5, and a drop in IgE synthesis, in mastocytes and in eosinophil activation. Both these components are features of the protective response against helminthes. Accordingly, there exists a higher prevalence of schistosomiasis and strongyloidiases in patients infected by HTLV-1.24 There may also be a dissemination of S. stercoralis with severe forms of strongyloidiasis.25
2.3. Human papilloma virus (HPV)
HPV is a DNA virus that, apart from causing verruca vulgaris and condylomata acuminata, is strongly associated with the development of cervical neoplasia and skin cancer, mainly in immunosuppressed individuals. HPV involvement with skin cancer was also shown in patients with epidermodysplasia verruciform in which viral DNA was detected in macular lesions.26
Immune response against HPV in general is mediated by cellular immune response, regardless of whether class IgG and IgA antibodies against antigenic fractions are found in the cervical mucous of patients with cervical neoplasia.27,28 Inflammatory infiltrate consisting of macrophages and CD4+ cells is observed in spontaneously regressing condylomata. The lymphoproliferative response of antigen-specific T CD4+ cells to E2 proved to be associated with the elimination of HPV. On the other hand, specific CD8+ cells for antigens E6 and E7 are found in patients with large lesions or a cervical tumor. Furthermore, type 1 response reduction with a low production of IL-2, IFN-gamma and TNF-alpha is observed in patients with a high-grade intraepithelial lesion.29
3. IMMUNE RESPONSE IN INFECTIONS CAUSED BY PROTOZOANS
The main diseases caused by protozoans in human beings are leishmaniases, Chagas disease, malaria, toxoplasmosis and amebiasis. Protozoans are infectious intracellular agents that usually infect the host for long periods of time, owing to mechanisms that allow them to evade from aggressions mediated by the immune system. In addition, infections by protozoans usually only cause disease in some infected patients. This indicates that in most cases the immune system does not allow large scale multiplication of protozoans or the infection to spread, though it is unable to foster sterilization. Accordingly, these agents may remain in the host for its entire lifespan even without causing disease, unless this balance is lost by immune depression or by precipitation of an exacerbated immunitary response with tissue inflammation.
Various immune response components participate in the protection mechanism against protozoans, but these microorganisms manage to evade this protection mechanism.30 Whereas in vitro the Leishmania promastigotes are highly sensitive to the complement, infectant forms resist their action. Tripanosoma cruzi, for instance, has a feature of preventing the complement's activation insofar it covers itself with the host's molecules as the degradation accelerator factor (DAF). Leishmania are also susceptible to the action of neutrophils, cells having a large potential to produce hydrogen peroxide and NO. But when penetrating the host, they infect the macrophages and make them vulnerable to a neutrophil attack. The adaptive response against protozoans occurs after the presentation of antigens by macrophages and dendritic cells, via MHC class II to the T cells. As other cells may be infected, and macrophages and dendritic cells also express MHC class I molecules, TCD8+ cells are also activated in protozoan infections. Chart 2 shows the immune protection mechanisms against some clinically important protozoans.
With the exception of Giardia lamblia, which may cause severe infection in patients who have an antibody production deficiency, immune cellular response is fundamental in protecting against infections caused by protozoans.
Whereas with infections caused by intracellular agents immune response deviated by the Th2 pole could incur damages, due notably to the fact that susceptibility to infection increases and this in turn allows the multiplication and dissemination of the parasite, the concept of whether a potent Th1 response is protective must be addressed with some skepticism. In various protozoan-caused diseases, there is evidence that an exacerbated immune response is involved in tissue damage: in amebiasis, it depends on neutrophil action; in Chagas disease it is mediated by CD4+ and CD8+ cells;32 a massive production of TNF-alpha and NO, documented in the pathogens of cerebral malaria.33 These facts indicate that a balanced performance of the immune system is very important in order to contain the parasite without incurring any tissue destruction, so that despite remaining in the host, the infecting agent does not cause disease to the human being.
The pathogenesis of diverse clinical forms of leishmaniasis exemplifies well the importance of Th1 response in the control and genesis of tissue lesions. The most common clinical forms of leishmaniasis are tegumentary leishmaniasis (cutaneous leishmaniasis, mucous leishmaniasis and diffuse cutaneous leishmaniasis) and visceral leishmaniasis. Chart 3 shows the association between diverse clinical forms of leishmaniasis, the Leishmaniasis species and immune response.
After inoculation of Leishmaniasis in the skin and the macrophage invasion, in individuals unable to produce IFN-gamma and activate macrophages, Leishmaniasis disseminates. Depending on the species, the latter causes visceral leishmaniasis (L. chagasi), or diffuse cutaneous leishmaniasis (L. amazonensis). In these patients, it is easy to understand the development of the disease, which occurs through IFN-gamma deficiency and high production of IL-10. Restoration of immune response in vitro in visceral leishmaniasis may be observed by neutralizing IL-10 or adding IL-12 to peripheral blood mononuclear cell cultures (PBMNCC).34
More atypical is what occurs to cutaneous leishmaniasis and mucous leishmaniasis, situations in which a strong Th1 deviation exists. Even though the number of parasites in the skin is thick or even absent, the lesion tends to develop. PBMNCC of individuals with cutaneous leishmaniasis and mucous leishmaniasis stimulated with the Leishmania antigen produces large amounts of IFN-gamma, IL-2 and only slight amounts of IL-10. As the immune system does not usually manage to completely destroy leishmania, this strong Th1 response prompts the occurrence of a very intense inflammatory reaction and damage to the tissues themselves. This results in the appearance of ulcers on the skin and mucosa. This damaged tissue also participates considerably on the accentuated production of TNF-alpha and NO. The evidence that cellular immune response participates in the pathogenesis of cutaneous leishmaniasis and mucous leishmaniasis includes: 1) premature treatment of the infection does not prevent the appearance of the lesion;35 2) the existence of a strong inflammatory reaction in the tissue with an increased expression of TNF-alpha, IFN-gamma and a few parasites on the lesion;34,36 3) association of an antimonial with an inhibitor TNF-alpha drug cures patients with mucous leishmaniasis, which are otherwise refractory to antimonial treatment.37
4. IMMUNE RESPONSE TO FUNGUS
The main protection mechanism against funguses is developed by phagocytes, which destroy them by producing NO and other components developed by these cells. In addition, there is participation of IFN-gamma. This enhances the function of neutrophils and macrophages, though there is no evidence of cytotoxic activity by T CD8+ cells. However, patients presenting with neutropenia (less than 500 neutrophils/mm3) or that have frequent cellular immune deficiency present with recurrent mycoses and occasionally develop severe and deep forms.38
Whereas a large number of fungus species may cause diseases in humans, the majority of them cause limited disease without greater clinical repercussions. Among the funguses associated with morbidity in Brazil, we can highlight Candida albicans, Criptococcus neoformas and Paraoccidiodis brasiliensis. In spite of the fact that infection by C. albicans regularly causes light infections with no greater consequences, HIV-positive patients not only present with a high prevalence of C. albicans infection, but esophagus, stomach, and intestine involvement are among the most recurrent infections. In children presenting alterations in cellular immune response and multiple endocrinal disturbances, a rare picture of chronic mucocutaneous candidiasis is described. In these children, one observes a reduction in Th1 response and severe cutaneous, mucous and ungual lesions.39
Despite the fact that vaginal candidiasis is extremely frequent but with no greater repercussions, roughly 5% of women at reproductive age do present with a condition of recurrent vaginal candidiasis due to the absence of or low levels of IFN-gamma, which may be restored in vitro by neutralizing IL-10.40 Although there is no documentation of Th2 response against C. albicans antigens, the high rate of atopia in these patients suggest that an immediate hypersensitivity reaction to diverse antigens may participate in the disease pathogenesis. Moreover, some cases may bring benefits to immunotherapy.41
Criptococcus neoformans may cause lung diseases and compromise the central nervous system in immunosuppressed patients. P. braziliensis is the causal agent of south-American blastomycosis. South-American blastomycosis is characterized by involvement of the ganglia, bucal mucosa and respiratory apparatus. In most infected patients, the agent is controlled and the individual remains completely asymptomatic. When there is no development of Th1, there is dissemination of the fungus with involvement of the organs of the reticuloendothelial and pulmonary system; in this context, the role of IL-4 seems important, given that in an experimental model the absence of this cytokine protects against severe pulmonary disease.42
5. IMMUNOLOGICAL RESPONSE IN HELMINTH INFECTIONS
The immune response mechanisms in helminth infections are manifold owing to the size and metabolic diversity of the parasites, which are antigenically complex. An additional problem is that the parasites may survive in the host for several years. As a result mechanisms are evaded, an example of which occurs with S. mansoni which ends up being covered by host antigens, and are no longer foreign for the immunological system.43
Although the complement and other factors of natural immune response might contribute to protecting against the helminth infections, specific immune response with the production of antibodies and cytokines is important. The T CD4+ or TCD8+ type 2 cells are producers of cytokines like IL-4, IL-5 and IL-13 which, among their other functions, induce IgE production by B cells and activate the fundamental components in protecting against helminths, namely eosinophils, mastocytes and basophiles, respectively. Class IgE antibodies bind to circulating basophiles or tissue mastocytes. This induces the release of histamine and other reaction mediators of immediate hypersensitivity leading to the destruction of helminths. The IgE produced in high levels in type Th2 immunological response has been related to protecting against reinfection by S. mansoni.45 Eosinophils also have the capacity to destroy schistosomula and strongyloides through the cytotoxicity cellular mechanism that depends on the antibody. Th2 type cells are associated with a resistance to infection not only by S. mansoni, but by intestinal helminths, for example S. stercoralis and A. lumbricoides. IL-4 stimulates IgE production and, in combination with IL-13, mastocytes. This results in increased secretion of inflammation mediators, secretion of mucus and enhanced contractility of intestinal musculature, which facilitates the expulsion of adult worms.47
In the acute phase of schistosomiasis, the clinical manifestations of fever, asthenia, weight loss, abdominal pain, diarrhea and coughing, in addition to complications such as pleuritis and pericarditis, result from the presence of TNF-alpha, IL-1 and IL-6, and also the deposition of immune complexes.48 Improvement of symptomatology coincides with the production of IL-10 induced by egg antigens in the chronic phase.49 This phase also features secretion of IL-4, IL-5 and IL-13,50,51 that experimental models participate in the formation of granuloma and hepatic fibrosis, and yet in the pathogenesis of schistosomiasis. 
REFERENCES
Received on November 15, 2004
Approved by the Editorial Council and accepted for publication on November 28, 2004
- 1. Janeway CA Jr. How the immune system protects the host from infection. Microbes Infect. 2001;3:1167-71.
- 2. Cooke A, Zaccone P, Raine T, Phillips JM, Dunne DW. Infection and autoimmunity: are we winning the war, only to lose the peace? Trends Parasitol. 2004;20:316-21.
- 3. Pasare C, Medzhitov R. Toll-like receptors and acquired immunity. Semin Immunol. 2004;16:23-6.
- 4. Debenedictis C, Joubeh S, Zhang G, Barria M, Ghohestani RF. Immune functions of the skin. Clin Dermatol. 2001; 19:573-85.
- 5. Romani N, Holzmann S, Tripp CH, Koch F, Stoitzner P. Langerhans cells - dendritic cells of the epidermis. APMIS. 2003;111:725-40.
- 6. Mosmann TR, Coffman RL. TH1 and TH2 cells: different patterns of lymphokine secretion lead to different functional properties. Annu Rev Immunol. 1989;7:145-73.
- 7. Mills KH, McGuirk P. Antigen-specific regulatory T cells-their induction and role in infection. Semin Immunol. 2004;16:107-17.
- 8. Britton WJ, Lockwood DNJ. Leprosy. The Lancet. 2004;363:1209-19.
- 9. Goulart IMB, Penna GO, Cunha G. Imunopatologia da hanseníase: a complexidade dos mecanismos da resposta imune do hospedeiro ao Mycobacterium leprae. Rev Bras Med Trop. 2002, 35:365-75.
- 10. Naafs B. Leprosy reactions. New knowledge. Trop Geogr Med. 1994;46:80-4.
- 11. Moreira AL, Sampaio EP, Zmuidzinas A, Frindt P, Smith KA, Kaplan G. Thalidomide exerts its inhibitory action on tumor necrosis factor alpha by enhancing mRNA degradation. J Exp Med. 1993;177:1675-80.
- 12. Sarno EN, Grau GE, Vieira LM, Nery JA. Serum levels of tumour necrosis factor-alpha and interleukin-1 beta during leprosy reactional states. Clin Exp Immunol. 1991;84:103-8.
- 13. Yamamura M, Wang XH, Ohmen JD, Uyemura K, Rea TH, Bloom BR, et al Cytokine patterns of immunologically mediated tissue damage. J Immunol. 1992;149:1470-5.
- 14. Foss NT. Aspectos imunológicos da hanseníase. Medicina (Ribeirão Preto) 1999; 30: 335-9.
- 15. Modlin RL, Melancon-Kaplan J, Young SMM, Pirmez C, Kino H, Convit J, et al Learning from lesions: patterns of tissue inflammation in leprosy. Proc Natl Acad Sci USA. 1988; 85: 1213-7.
- 16. Barrington R, Zhang M, Fischer M, Carroll MC. The role of complement in inflammation and adaptive immunity. Immunol Rev. 2001;180:5-15.
- 17. Dong VM, McDermott DH, Abdi R. Chemokines and diseases. Eur J Dermatol. 2003;13:224-30.
- 18. Manetti R, Parronchi P, Giudizi MG, Piccinni MP, Maggi E, Trinchieri G, et al Natural killer cell stimulatory factor (interleukin 12 [IL-12]) induces T helper type 1 (Th1)-specific immune responses and inhibits the development of IL-4-producing Th cells. J Exp Med. 1993;177:1199-204.
- 19. Bacharier LB, Geha RS. Molecular mechanisms of IgE regulation. J Allergy Clin Immunol. 2000;105:S547-58.
- 20. Caille V, Bossi P, Grimaldi D, Vieillard-Baro A. Physiopathology of severe sepsis. Presse Med. 2004;33:256-61.
- 21. Chadha KC, Ambrus JL Jr, Dembinski W, Ambrus JL Sr. Interferons and interferon inhibitory activity in disease and therapy. Exp Biol Med. 2004; 229:285-90.
- 22. Kedzierska K, Crowe SM. Cytokines and HIV-1: interactions and clinical implications. Antivir Chem Chemother. 2001;12:133-50.
- 23. Barmak K, Harhaj EW, Wigdahl B. Mediators of central nervous system damage during the progression of human T-cell leukemia type I-associated myelopathy/tropical spastic paraparesis. J Neurovirol. 2003;9:522-9.
- 24. Porto AF, Neva FA, Bittencourt H, Lisboa W, Thompson R, Alcântara L, Carvalho EM. HTLV-1 decreases Th2 type of immune response in patients with strongyloidiases. Parasite Imunol. 2001; 23:503-07.
- 25. Porto MA, Muniz A, Oliveira Junior J, Carvalho EM. Clinical and immunological consequences of the association between HTLV-1 and strongyloidiasis. Rev Soc Bras Med Trop. 2002;35:641-9.
- 26. Pfister H, Ter Schegget J. Role of HPV in cutaneous premalignant and malignant tumors. Clin Dermatol. 1997;15:335-47.
- 27. Dillner L, Bekassy Z, Jonsson N, Moreno-Lopez J, Blomberg J. Detection of IgA antibodies against human papillomavirus in cervical secretions from patients with cervical intraepithelial neoplasia. Int J Cancer. 1989;43:36-40.
- 28. Dillner L, Fredriksson A, Persson E, Forslund O, Hansson BG, Dillner J. Antibodies against papillomavirus antigens in cervical secretions from condyloma patients. J Clin Microbiol. 1993;31:192-7.
- 29. Lee BN, Follen M, Shen DY, Malpica A, Adler-Storthz K, Shearer WT, et al. Depressed type 1 cytokine synthesis by superantigen-activated CD4+ T cells of women with human papillomavirus-related high-grade squamous intraepithelial lesions. Clin Diagn Lab Immunol. 2004;11:239-44.
- 30. Zambrano-Villa S, Rosales-Borjas D, Carrero JC, Ortiz-Ortiz L. How protozoan parasites evade the immune response. Trends Parasitol. 2002;18:272-8.
- 31. Ventura-Juarez J, Jarillo-Luna RA, Fuentes-Aguilar E, Pineda-Vazquez A, Munoz-Fernandez L, Madrid-Reyes JI, et al Human amoebic hepatic abscess: in situ interactions between trophozoites, macrophages, neutrophils and T cells. Parasite Immunol 2003;25:503-11.
- 32. Gomes JA, Bahia-Oliveira LM, Rocha MO, Martins-Filho OA, Gazzinelli G, Correa-Oliveira R. Evidence that development of severe cardiomyopathy in human Chagas' disease is due to a Th1-specific immune response. Infect Immun. 2003;71:1185-93.
- 33. Artavanis-Tsakonas K, Tongren JE, Riley EM. The war between the malaria parasite and the immune system: immunity, immunoregulation and immunopathology. Clin Exp Immunol. 2003;133:145-52.
- 34. Ribeiro de Jesus A, Almeida RP, Lessa H, Bacellar O, Carvalho EM. Cytokine profile and pathology in human leishmaniasis. Braz. J. Med. Biol. Res. 1998;31:143-8.
- 35. Machado P, Araujo C, Da Silva AT, Almeida RP, D'Oliveira Jr A, Bittencourt A, et al Failure of early treatment of cutaneous leishmaniasis in preventing the development of an ulcer. Clin Infect Dis. 2002;34:E69-73.
- 36. Pirmez C, Yamamura M, Uyemura K, Paes-Oliveira M, Conceição-Silva F, Modlin RL. Cytokine patterns in the pathogenesis of human leishmaniasis. J Clin Invest. 1993; 91: 1390-5.
- 37. Lessa HA, Machado P, Lima F, Cruz AA, Bacellar O, Guerreiro J, et al Successful treatment of refractory mucosal leishmaniasis with pentoxifylline plus antimony. Am J Trop Med Hyg. 2001; 65:87-9.
- 38. Romani L. Immunity to fungal infections. Nat Rev Immunol. 2004;4:1-23.
- 39. Lilic D. New perspectives on the immunology of chronic mucocutaneous candidiasis. Curr Opin Infect Dis. 2002;15:143-7.
- 40. Carvalho LP, Bacellar O, Neves N, de Jesus AR, Carvalho EM. Downregulation of IFN-gamma production in patients with recurrent vaginal candidiasis. J Allergy Clin Immunol. 2002;109:102-5.
- 41. Moraes PS, de Lima Goiaba S, Taketomi EA. Candida albicans allergen immunotherapy in recurrent vaginal candidiasis. J Investig Allergol Clin Immunol. 2000;10:305-9.
- 42. Pina A, Valente-Ferreira RC, Molinari-Madlum EE, Vaz CA, Keller AC, Calich VL. Absence of interleukin-4 determines less severe pulmonary paracoccidioidomycosis associated with impaired Th2 response. Infect Immun. 2004;72:2369-78.
- 43. Neva FA, Brown HW. Basic clinical parasitology. Appleton & Lange; 1994. p. 107-144.
- 44. Else KJ, Finkelman FD. Intestinal nematode parasites, cytokines and effector mechanisms. Int J Parasitol. 1998; 28: 1145-58.
- 45. Demeure CE, Rihet P, Abel L, Ouattara M, Bourgois A, Dessein AJ. Resistance to Schistosoma mansoni in humans: influence of the IgE/IgG4 balance and IgG2 in immunity to reinfection after chemotherapy. J Infect Dis. 1993; 168: 1000-8.
- 46. MacDonald A, Araujo MI, Pearce EJ. Immunology of parsitic helminth infection. Infect Immun. 2002; 70:427-33.
- 47. Finkelman FD, Shea-Donohue T, Goldhill J, Sullivan CA, Morris SC, Madden KB, Gause WC, Urban JF Jr. Cytokine regulation of host defense against parasitic gastrointestinal nematodes: lessons from studies with rodent models. Annu Rev Immunol.1997; 15:505-33.
- 48. Butterworth AE. Immunological aspects of human schistosomiasis. Br Med Bull. 1998; 54:357-68.
- 49. Grzych JM, Pearce E, Cheever A, Caulada ZA, Caspar P, Heiny S, et al Egg deposition is the major stimulus for the production of Th2 cytokines in murine schistosomiasis mansoni. J Immunol. 1991; 146: 1322-7.
- 50. Araujo MI, de Jesus AR, Bacellar O, Sabin E, Pearce E, Carvalho EM. Evidence of a T helper type 2 activation in human schistosomiasis. Eur J Immunol. 1996; 26: 1399-403.
- 51. Malaquias LC, Falcão PL, Silveira AM, Gazzinelli G, Prata A, Coffman RL, et al Cytokine regulation of human immune response to Schistosoma mansoni: analysis of the role of IL-4, IL-5 and IL-10 on peripheral blood mononuclear cell responses. Scand J Immunol. 1997; 46: 393-8.
Publication Dates
-
Publication in this collection
30 Mar 2005 -
Date of issue
Dec 2004
History
-
Accepted
28 Nov 2004 -
Received
15 Nov 2004