Abstracts
The entomophatogenic bacterium Bacillus thuringiensis produces crystal proteins, named Cry proteins which are encoded by the cry genes. This bacterium is used on biological control of important economical pests, as well as in the control of disease´s vectors, such as Aedes aegypti, a mosquito that transmits the dengue viruses. Isolates of this bacterium can be characterized by the content of cry genes and this prediction helps target different insect orders. In this research, we isolated 76 colonies of B. thuringiensis from 30 soil samples that were taken from Ilha Bela (SP, Brazil), a place where simulids are already biologically controlled by B. thuringiensis, to find bacterial isolates that were capable of controlling A. aegypti. The 16S ribosomal subunit genes of the selected isolates were sequenced, and the isolates were molecularly characterized based on their Dipteran-specific cry gene contents. Eight of the 76 isolates (10.52%) contained the cry4Aa, cry4Ba or cry10Aa genes, these isolates were carried out against A. aegypti larvae on bioassay. The presence or absence of specific cry genes was associated with the observed average larval mortalities. From the 76 isolates, seven (9.2%) were potentially able to control A. aegypti larvae. Therefore these are promising isolates for the biological control of A. aegypti larvae.
dengue; biological control; Dipteran; 16S rRNA
Bacillus thuringiensis é entomopatogênica, por produzir proteínas cristais, denominadas proteínas Cry, as quais são codificadas pelos genes cry. Essa bactéria atua no controle biológico de insetos-praga de culturas economicamente importantes, bem como no controle de insetos vetores causadores de doenças, como o Aedes aegypti, mosquito transmissor do vírus da dengue. Os isolados dessa bactéria podem ser caracterizados pelo conteúdo de genes cry que possuem e, assim, predizer o alvo de controle dos mesmos às diferentes ordens de insetos. Com o objetivo de encontrar isolados eficientes no controle do vetor A. aegypti, o presente trabalho isolou 76 colônias de B. thuringiensis a partir de 30 amostras de solo oriundas de Ilhabela-SP, município que se caracteriza por realizar controle biológico de simulídeos com essa bactéria. Os 76 isolados foram sequenciados na região da subunidade ribossomal 16S e caracterizados molecularmente quanto ao conteúdo de genes cry díptero-específicos. No total, oito isolados (10,52% do total) apresentaram bandas para os genes cry4Aa, cry4Ba e cry10Aa, sendo os mesmos testados contra larvas de A. aegypti por meio de bioensaios. A presença e/ou ausência dos genes cry foi associada à mortalidade média de larvas. Dentre os isolados estudados, sete (9,2% do total) apresentaram elevado potencial de controle às larvas de A. aegypti, sendo assim considerados como promissores para o manejo do controle biológico de larvas de A. aegypti com a bactéria B. thuringiensis.
dengue; controle biologico; Dipterous; rRNA 16S
ECOLOGY
Isolation of Bacillus thuringiensis strains that contain Dipteran-specific cry genes from Ilha Bela (São Paulo, Brazil) soil samples
Isolamento de linhagens de Bacillus thuringiensis contendo genes cry díptero-específicos a partir de amostras de solo oriundas de Ilha Bela-São Paulo, Brasil
Campanini, EB.; Davolos, CC.; Alves, ECC.; Lemos, MVF. * * e-mail: mvictor@fcav.unesp.br
Laboratório de Genética de Bactérias e Biotecnologia Aplicada, Faculdade de Ciências Agrárias e Veterinária, Universidade Estadual Paulista - UNESP, Via de Acesso Prof. Paulo Donato Castellane, CEP 14884-900, Jaboticabal, SP, Brazil
ABSTRACT
The entomophatogenic bacterium Bacillus thuringiensis produces crystal proteins, named Cry proteins which are encoded by the cry genes. This bacterium is used on biological control of important economical pests, as well as in the control of disease´s vectors, such as Aedes aegypti, a mosquito that transmits the dengue viruses. Isolates of this bacterium can be characterized by the content of cry genes and this prediction helps target different insect orders. In this research, we isolated 76 colonies of B. thuringiensis from 30 soil samples that were taken from Ilha Bela (SP, Brazil), a place where simulids are already biologically controlled by B. thuringiensis, to find bacterial isolates that were capable of controlling A. aegypti. The 16S ribosomal subunit genes of the selected isolates were sequenced, and the isolates were molecularly characterized based on their Dipteran-specific cry gene contents. Eight of the 76 isolates (10.52%) contained the cry4Aa, cry4Ba or cry10Aa genes, these isolates were carried out against A. aegypti larvae on bioassay. The presence or absence of specific cry genes was associated with the observed average larval mortalities. From the 76 isolates, seven (9.2%) were potentially able to control A. aegypti larvae. Therefore these are promising isolates for the biological control of A. aegypti larvae.
Keywords: dengue, biological control, Dipteran, 16S rRNA.
RESUMO
Bacillus thuringiensis é entomopatogênica, por produzir proteínas cristais, denominadas proteínas Cry, as quais são codificadas pelos genes cry. Essa bactéria atua no controle biológico de insetos-praga de culturas economicamente importantes, bem como no controle de insetos vetores causadores de doenças, como o Aedes aegypti, mosquito transmissor do vírus da dengue. Os isolados dessa bactéria podem ser caracterizados pelo conteúdo de genes cry que possuem e, assim, predizer o alvo de controle dos mesmos às diferentes ordens de insetos. Com o objetivo de encontrar isolados eficientes no controle do vetor A. aegypti, o presente trabalho isolou 76 colônias de B. thuringiensis a partir de 30 amostras de solo oriundas de Ilhabela-SP, município que se caracteriza por realizar controle biológico de simulídeos com essa bactéria. Os 76 isolados foram sequenciados na região da subunidade ribossomal 16S e caracterizados molecularmente quanto ao conteúdo de genes cry díptero-específicos. No total, oito isolados (10,52% do total) apresentaram bandas para os genes cry4Aa, cry4Ba e cry10Aa, sendo os mesmos testados contra larvas de A. aegypti por meio de bioensaios. A presença e/ou ausência dos genes cry foi associada à mortalidade média de larvas. Dentre os isolados estudados, sete (9,2% do total) apresentaram elevado potencial de controle às larvas de A. aegypti, sendo assim considerados como promissores para o manejo do controle biológico de larvas de A. aegypti com a bactéria B. thuringiensis.
Palavras-chave: dengue, controle biologico, Dipterous, rRNA 16S.
1. Introduction
Dengue fever is a serious infectious disease that can be lethal and is caused by four serotypes of a flavivirus. The dengue virus is transmitted by the insect vector, Aedes aegypti (Diptera: Culicidae) (Linnaeus, 1762), and is mainly found in tropical and subtropical regions where the environmental conditions are favorable for the development and proliferation of the vector (Forattini, 1992). Despite numerous efforts, an effective vaccine against the dengue virus has not been developed (World Health Organization) (WHO, 1996). Thus, the insect vector is the only component of the epidemiological chain that can be targeted to prevent the transmission of the virus.
Initially, A. aegypti control strategies include the elimination of larval breeding sites and the administration of chemical insecticides to control adult insects. Unfortunately when insecticides were used to control A. aegypti, environmental impacts of the insecticides and the development of resistance were observed in Brazil (Macoris et al., 1999) and abroad (World Health Organization) (WHO, 1992), which generated increasing efforts to find more environmentally- friendly alternatives that would efficiently control the insect vector.
An alternative is the use of Bacillus thuringiensis (Eubacteria: Bacillaceae) (Berliner, 1911) as a biological control agent. This entomopathogenic bacterium produces crystal proteins, which were are encoded by specific cry genes and control this insect disease vector. The Cry proteins are advantageous biological control agents because they are extremely toxic to specific target insects and are not harmful to humans, plants or animals that are present in the same environments (De-Souza et al., 1999). This characteristic is important for dengue control strategies because the control agents need to be applied in urban areas where the mosquito vectors primarily reside.
Currently, B. thuringiensis strains are used as biological control agents. Standard strains of B. thuringiensis var. israelensis (T 14001), which produce specific proteins that are toxic to Dipterans, have been successfully used to control Dipterans in Southern Brazil (Rio Grande do Sul state) (Ruas-Neto, 1984) on the coast of the São Paulo state in Serra do Mar (Araujo-Coutinho, 1995).
In this study, we select and characterize Dipteran-specific B. thuringiensis isolates from soil samples of Ilha Bela (São Paulo, Brazil) by molecular analyses. Furthermore, we determined if any of isolates showed biological control characteristics against A. aegypti larvae by performing bioassays and correlating the presence or absence of specific cry genes with the observed larval mortalities. Efficacies of isolates were compared to the efficacy of the standard B. thuringiensis var. israelensis (T 14001) isolate.
2. Material and Methods
2.1. Obtaining bacterial isolates
Bacterial colonies were isolated from 30 soil samples (1 g each) that were collected from coastal cities of Ilha Bela (São Paulo, Brazil) (location: 23° 13' 56" S and 45° 42' 83" W). During October and November of 2005. To isolate the colonies, 10 mL of Milli-Q sterile water were added to the samples and after being agitated were allowed to sit for 1 hour to allow for sample sedimentation. The solutions were carefully filtered, the filtrates were heated at 80 °C for 10 minutes to eliminate any unsporulated microorganisms and the filtrates were diluted 10-fold and 100-fold. One hundred microliters of each dilution were inoculated onto Petri dishes containing NA (agar nutrient) medium (0.5% yeast extract, 0.1% tryptone, 0.17 M NaCl and 0.15% bacteriological agar).
Plates were incubated in a BOD at 30 °C for 36 hours, and morphologies of the colonies that grew were assessed. Colonies that showed bacilli features were selected and microscopically observed to verify the presence of spores and crystals. The selected colonies were then inoculated on NA medium and incubated at 30 °C for 5 days to allow sporulation. After sporulation, 300 µL of Milli-Q water was added to the medium plate and the isolates suspension (water and spores) were placed on sterile filter paper strips and stored at 10 °C in cryogenic tubes. These isolates belong to the LGBBA collection which is located at São Paulo State University 'Julio de Mesquita Filho', Jaboticabal-SP.
2.2. DNA extractions and sequencing of the 16S ribosomal subunit genes
The total DNA from isolates was obtained by using the InstaGene Matrix ion exchange resin (Bio-Rad). The DNA samples were sequenced using an oligonucleotide primer pair that amplified the 16S ribosomal subunit gene (Kurokawa, et al., 1997). The sequencing reactions were performed using the DNA Sequencing-Big Dye Terminator Cycle Sequencing Ready ABI Prism reaction kit, version 3, and the ABI 3100 (Applied Biosystems) in a capillary system.
2.3. Molecular characterizations by polymerase chain reactions (PCR)
Dipteran-specific genes were amplified by PCR using the protocols and oligonucleotide primers (cry4Aa, cry4Ba, and cry10Aa) that had been previously described by Costa (2010). DNA from the standard B. thuringiensis var. israelensis (T 14001) strain was used as a positive control, while DNA from the standard B. thuringiensis var. kurstaki-HD1 strain was used as a negative control. Sterile water was used as the template instead of genomic DNA as an additional negative control.
2.4. Bioassays with A. aegypti larvae
Dipteran-specific isolates detected by PCR analyses were tested for activity against A. aegypti larvae using bioassays according to Costa (2010). Twenty larvae on third instar were layed on plastic cups (300 mL) with spore suspension and water. Average mortalities were calculated and subjected to variance analyses by the F-test. Means were compared using Tukey's test and the isolates activity were considered significant at 1% of probability.
3. Results
Seventy-six different colonies of B. thuringiensis, which were selected based on their morphological characteristics that were previously defined by Sosa-Gómez et al. (1998), were isolated from 30 soil samples. When cry genes were PCR amplified from the isolates, 2 of the isolates (2.63% of the entire collection) contained cry4Aa, 8 of the isolates (10.52% of the entire collection) contained cry4Ba and 3 of the isolates (3.95% of the entire collection) contained cry10Aa. Specifically, isolates IB17.2, IB17.4, IB18.1, IB20.1, IB20.3, IB20.5, IB20.6 and IB23.2 had cry4Ba, isolates IB17.2, IB17.4 and IB18.1 had cry10Aa, and isolates IB17.2 and IB18.1 had cry4Aa and all of other tested genes. The cry4Aa gene occurred at the lowest frequency in the population.
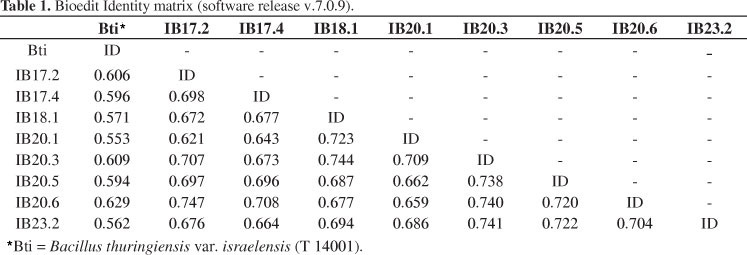
Sequences of the 16S ribosomal subunit genes from the isolates mentioned above and from the positive control, B. thuringiensis var. israelensis (T 14001), were aligned using the ClustalW tool (Thompson et al., 1997). An identity matrix was then created using the Bioedit software v.7.0.9 (Hall, 1999) (Table 1). Results indicated that all isolates were uniquely different from each other and from the control strain B. thuringiensis var. israelensis (T 14001). The most divergence was found between the control strain and the isolate IB20.1 (55.3% of identity). All of the Dipteran-specific isolates were toxic to A. aegypti larvae, and most of the isolates caused 100% larval mortality after 10 minutes to exposure of the larvae to the bacteria suspension. Isolates IB20.5 and IB23.2 were the least effective of these isolates against larvae and caused 100% larval mortality after 2 and 4 hourss of inoculation, respectively.
The analyses of mortality (Table 2) revealed that only isolate IB23.2 showed marginal control (71.66%) of A. aegypti larvae when compared to the other seven isolates, which presented a high control (>75%) of the larvae. In fact, isolate IB20.5 had an average larval mortality of 96.25%, while isolates IB17.2, IB17.4, IB18.1, IB20.1, IB20.3 and IB20.6 had average larval mortalities of 100%, which were comparable to the positive control, B. thuringiensis var. israelensis (T 14001).
4. Discussion
In all of the soil samples that were collected in this study, bacterial growth was detected with an average of 2.53 B. thuringiensis isolates/sample, which corresponds with the averages of 3.45, 2.35 and 1.91 B. thuringiensis isolates/sample that were previously described by Polanczyk et al. (2003), Valicente and Barreto (2003), and Barreto (2005), respectively. Because the isolates were initially characterized from the soil samples by morphological features, we sequenced the 16S ribosomal subunit genes of the isolates to confirm that they were B. thuringiensis. The sequences were compared using BLASTN, and the results indicated that all of the isolates displayed similarity above 93% to the B. thuringiensis strain DTAG2 16S ribosomal RNA gene, partial sequence (accession number: EF113701.1) or Bacillus thuringiensis strain CESNVD 10 16S ribosomal RNA gene, partial sequence (accession number: FJ649680.1).
We amplified Dipteran-toxic cry genes from eight distinct isolates, which represent approximately 10.52% of the group that was analyzed. This percentage is slightly higher than the percentage that was previously reported by Bravo et al. (1998). The authors reported that 7.9% of the strains that were isolated contained genes that encoded Dipteran-toxic proteins. Of the eight isolates that we identified, seven were highly effective against A. aegypti in larval bioassays. According to Dias et al. (1992), only strains of the B. thuringiensis var. israelensis (T 14001) lineage effectively controlled Dipteran larvae. However, this study demonstrates that additional B. thuringiensis isolates have the abilities to control Dipteran larvae as effectively as strains of the B. thuringiensis var. israelensis (T 14001) lineage. The effectiveness of the isolates in this study might be related to the constant introduction of the B. thuringiensis var. israelensis (T 14001) program used by the Ilha Bela's city health authorities.
The use of B. thuringiensis as a safe and efficient biological control agent requires continued isolation. The seven new B. thuringiensis isolates that were found in this study, which all efficiently control A. aegypti larvae, may be potential alternative biological control agents of this insect vector because they provide the same insecticidal activity as the commonly used B. thuringiensis lineage, B. thuringiensis var. israelensis (T 14001), and represent a viable option to minimize the emergence of possible Cry-resistant insects.
Effective A. aegypti control strategies are urgently needed and extremely important. Because chemical insecticides are becoming less efficacious with the emergence of resistant strains and present detrimental environmental impacts, biological control strategies that are effectively applied can provide the desired efficacy against specific insect vectors and consequently decrease the spread of dengue fever throughout Brazil and the world.
Acknowledgements - The first author is grateful to the São Paulo State Research Foundation (FAPESP, Process # 06/58080-6) for granting an undergraduate fellowship to do this work. The authors are also grateful to the Laboratório de Ecologia Química de Insetos Vetores, Departamento de Parasitologia da Universidade Federal de Minas Gerais (UFMG) - Instituto de Ciências Biológicas (ICB) for supplying the larvae used in the bioassay.
Received February 04, 2011
Accepted July 29, 2011
Distributed May 31, 2012
- ARAUJO-COUTINHO, CJPC., 1995. Biological control program against simuliids in the Stade of São Paulo, Brasil. Memórias do Instituto Oswaldo Cruz, vol. 90, p. 131-133.
- BARRETO, MR., 2005. Prospecção e caracterização de genes de Bacillus thuringiensis e determinação de sua atividade entomocida para espécies de Lepidoptera e Coleoptera pragas da soja Curitiba: Universidade Federal do Paraná. 107 p. Tese de Doutorado em Ciências Biológicas.
- BRAVO, A., SARABIA, S., LOPEZ, L., ABARCA, C., ORTIZ, A., ORTIZ, M., LINA, L., VILLALOBOS, FJ., PEÑA, G., NUÑEZ-VALDEZ, ME., SOBERÓN, M. and QUINTERO, R., 1998. Characterization of cry genes in a mexican Bacillus thuringiensis strain collection. Applied and Environmental Microbiology, vol. 64, p. 4965-4972.
- COSTA, JRV., ROSSI, JR., MARUCCI, SC., ALVES, ECC., VOLPE, HXL., FERRAUDO, AS., LEMOS, MVF. and DESIDÉRIO, JA., 2010. Atividade tóxica de isolados de Bacillus thuringiensis a larvas de Aedes Aegypti (l.) (Diptera: Culicidae). Neotropical Entomology, vol. 39, p. 757-766. PMid:21120386. http://dx.doi.org/10.1590/S1519-566X2010000500015
- DE-SOUZA, MT., LIMA, MI., SILVA-WERNECK, JO., DIAS, JCS. and RIBEIRO, BM., 1999. Ultrastructural and molecular characterizat ion of the parasporal Crystal proteins of Bacillus thuringiensis subsp. kusrtaki S93 active against Spodoptera frugiperda Biocell, vol. 23.1, p. 43-49.
- DIAS, JMCS., 1992. Produção e utilização de bioinseticidas bacterianos. Pesquisa Agropecuária Brasileira, vol. 27, no. 1, p. 59-76.
- FORATTINI, OP., 1992. Ecologia, epidemiologia e sociedade. São Paulo: EdUSP; Livraria Editora Artes Médicas Ltda. 271 p.
- HALL, TA., 1999 BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series, vol. 41, p. 95-98.
- KUROKAWA, K., TANI, K. and NASU, M., 1997. Direct in situ PCR method for the detection of verotoxin-producing Escherichia coli Japanese Journal of Bacteriology, vol. 52, p. 513-518. PMid:9155208. http://dx.doi.org/10.3412/jsb.52.513
- MACORIS, MLG., ANGRIGHETTI, MTM., GLASSER, CM., GARBELOTI, VC. and CIRINO, VCB., 1999. Alteração da resposta de suscetibilidade de Aedes aegypti a inseticidas organofosforados em municípios do Estado de São Paulo. Revista da Saúde Pública, vol. 33, p. 521-522. PMid:10576756.
- POLANCZYK, RA., 2003. Isolamento de Bacillus thuringiensis berliner a partir de amostras de solo e sua patogenecidade para Spodoptera frugiperda (J.E.SMITH) (Lepidoptera: Noctuidae) Piracicaba: Escola Superior de Agronomia, Universidade de São Paulo. 158 p. Tese de Doutorado em Fitotecnia.
- RUAS-NETO, AL., 1984. Bacillus thuringiensis var. israelensis (T 14001) como alternativa no controle de simulídeos no Rio Grande do Sul suscetibilidade a campo. B Saúde, vol. 11, p. 21-26.
- SOSA-GÓMEZ, DR., TIGANO, MS. and ALVES, SB., 1998. Caracterização de entomopatógenos. In ALVES, SB. (Ed.). Controle microbiano de insetos Piracicaba: FEALQ. p. 731-764.
- THOMPSON, JD., HIGGINS, DG. and GIBSON, TJ., 1997. ClustaW: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Research, vol. 22, p. 4673-4680.
- VALICENTE, FH and BARRETO, MR., 2003. Bacillus thuringiensis survey in Brazil: geographical distribution and insecticidal activity against Spodoptera frugiperda (J.E.Smith) (Lepidoptera: Noctuidae). Neotropical Entomology, vol. 32, no. 4, p. 639-644.
- World Health Organization - WHO, 1992. Vector resistance to pesticides Geneva: WHO.
- World Health Organization - WHO, 1996. Global Programme for Vacines and Imunization Vaccine: Research & Development. Geneva: WHO.
Publication Dates
-
Publication in this collection
21 June 2012 -
Date of issue
May 2012
History
-
Received
04 Feb 2011 -
Accepted
29 July 2011