Abstracts
The refugial hypothesis is treated as the definitive history of Amazonian forests in many texts. Surprisingly, this important theoretical framework has not been based on paleoecological data. Consequently, a model of Amazonian aridity during the northern hemispheric glaciation has been accepted uncritically. Ironically, the Refuge Hypothesis has not been tested by paleobotanical data. We present a revision of the concept of Neotropical Pleistocene Forest Refuges and test it in the light of paleocological studies derived from pollen analysis of Amazonian lake sediments deposited during the last 20,000 years. Our analysis is based primarily on paleoenvironmental data obtained from sites in Brazil and Ecuador. These data are contrasted with those that favor the hypothesis of fragmented tropical forests in a landscape dominated mainly by tropical savannas under an arid climate. The Ecuadorian data set strongly suggests a 5ºC cooling and presence of humid forests at the foot of the Andes, during the last Ice Age. The same climatic and vegetational scenario was found in the western Brazilian Amazon. On the other hand, somewhat drier conditions were observed in the central Amazon, but the landscape remained a forested landscape during the supposedly arid phases of the Late Quaternary. Data obtained from the Amazon Fan sediments containing pollen derived from extensive sections of the Amazon Basin, were fundamental to the conclusion that the predominance of savannas in this region is not supported by botanical data. Our revision of the assumptions derived from the Refuge Hypothesis indicates that it has succumbed to the test now permitted by a larger paleocological data set, which were not available during the golden age of this paradigm, when indirect evidence was considered satisfactory to support it.
Amazonia; refugia; fossil pollen; glaciations; Pleistocene; Holocene; Miocene; phylogeny; speciation diversity
A Hipótese dos Refúgios Florestais do Pleistoceno é aceita como a história definitiva da Amazônia por muitos autores. Surpreendentemente, este importante conceito não foi baseado em dados paleoecológicos. Como conseqüência, hoje temos um modelo teórico intimamente conectado à visão de uma Amazônia árida, durante o período de maior atividade glacial no hemisfério norte. Ironicamente, a Hipótese dos Refúgios não foi testada por dados paleobotânicos. Por isso, apresentamos uma revisão desse conceito e testamos a sua validade frente aos estudos paleoecológicos, derivados de análises palinológicas de sedimentos lacustres da Amazônia, depositados nos últimos 20.000 anos. Nossa análise baseia-se, principalmente, em dados paleoambientais obtidos em regiões da Amazônia do Brasil e do Ecuador, os quais são contrastados com informações que apóiam a hipótese da fragmentação florestal amazônica em uma paisagem dominada por savanas, sob clima predominantemente árido. Os dados do Ecuador sugerem enfaticamente um esfriamento da ordem de 5ºC e presença de florestas nos sopés dos Andes, durante a última glaciação. Este mesmo cenário climático e vegetacional foi encontrado na Amazônia Oriental Brasileira. Por outro lado, condições relativamente mais secas foram detectadas na Amazônia Central, mas ainda sugerem uma paisagem florestada durante as fases supostamente mais secas do Quaternário Tardio. Dados obtidos nos sedimentos do delta do Rio Amazonas, que contém pólen oriundo de extensas áreas da bacia, foram fundamentais à conclusão que a hipótese da predominância de savanas nessa região não tem apoio em dados botânicos. Nossa revisão das expectativas derivadas da Hipótese dos Refúgios indica que o modelo sucumbiu frente ao teste permitido por um banco de dados paleoecológicos, o qual não estava disponível durante a "idade dourada" deste paradigma, quando evidências indiretas eram consideradas satisfatórias para mantê-lo.
Amazônia; refúgios; pólen fóssil; glaciação; Pleistoceno; Holoceno; Mioceno; filogenia; diversidade e especiação
POINTS OF VIEW
The rise and fall of the Refugial Hypothesis of Amazonian speciation: a paleoecological perspective
Apogeu e declínio da Hipótese dos Refúgios para a especiação na Amazônia: uma perspectiva paleoecológica
Mark B. BushI; Paulo E. de OliveiraII
IDepartment of Biological Sciences, Florida Institute of Technology, Melbourne, Fl 32901, USA. e-mail: mbush@fit.edu; website: http://research.fit.edu/bushlab/
IILaboratório de Geociências, Universidade Guarulhos, Guarulhos, São Paulo 07023-070,Brazil.e-mail: paulo@bjd.com.br
ABSTRACT
The refugial hypothesis is treated as the definitive history of Amazonian forests in many texts. Surprisingly, this important theoretical framework has not been based on paleoecological data. Consequently, a model of Amazonian aridity during the northern hemispheric glaciation has been accepted uncritically.
Ironically, the Refuge Hypothesis has not been tested by paleobotanical data. We present a revision of the concept of Neotropical Pleistocene Forest Refuges and test it in the light of paleocological studies derived from pollen analysis of Amazonian lake sediments deposited during the last 20,000 years. Our analysis is based primarily on paleoenvironmental data obtained from sites in Brazil and Ecuador. These data are contrasted with those that favor the hypothesis of fragmented tropical forests in a landscape dominated mainly by tropical savannas under an arid climate.
The Ecuadorian data set strongly suggests a 5ºC cooling and presence of humid forests at the foot of the Andes, during the last Ice Age. The same climatic and vegetational scenario was found in the western Brazilian Amazon. On the other hand, somewhat drier conditions were observed in the central Amazon, but the landscape remained a forested landscape during the supposedly arid phases of the Late Quaternary. Data obtained from the Amazon Fan sediments containing pollen derived from extensive sections of the Amazon Basin, were fundamental to the conclusion that the predominance of savannas in this region is not supported by botanical data.
Our revision of the assumptions derived from the Refuge Hypothesis indicates that it has succumbed to the test now permitted by a larger paleocological data set, which were not available during the golden age of this paradigm, when indirect evidence was considered satisfactory to support it.
Key words: Amazonia, refugia, fossil pollen, glaciations, Pleistocene, Holocene, Miocene, phylogeny, speciation diversity
RESUMO
A Hipótese dos Refúgios Florestais do Pleistoceno é aceita como a história definitiva da Amazônia por muitos autores. Surpreendentemente, este importante conceito não foi baseado em dados paleoecológicos. Como conseqüência, hoje temos um modelo teórico intimamente conectado à visão de uma Amazônia árida, durante o período de maior atividade glacial no hemisfério norte.
Ironicamente, a Hipótese dos Refúgios não foi testada por dados paleobotânicos. Por isso, apresentamos uma revisão desse conceito e testamos a sua validade frente aos estudos paleoecológicos, derivados de análises palinológicas de sedimentos lacustres da Amazônia, depositados nos últimos 20.000 anos.
Nossa análise baseia-se, principalmente, em dados paleoambientais obtidos em regiões da Amazônia do Brasil e do Ecuador, os quais são contrastados com informações que apóiam a hipótese da fragmentação florestal amazônica em uma paisagem dominada por savanas, sob clima predominantemente árido.
Os dados do Ecuador sugerem enfaticamente um esfriamento da ordem de 5ºC e presença de florestas nos sopés dos Andes, durante a última glaciação. Este mesmo cenário climático e vegetacional foi encontrado na Amazônia Oriental Brasileira. Por outro lado, condições relativamente mais secas foram detectadas na Amazônia Central, mas ainda sugerem uma paisagem florestada durante as fases supostamente mais secas do Quaternário Tardio.
Dados obtidos nos sedimentos do delta do Rio Amazonas, que contém pólen oriundo de extensas áreas da bacia, foram fundamentais à conclusão que a hipótese da predominância de savanas nessa região não tem apoio em dados botânicos.
Nossa revisão das expectativas derivadas da Hipótese dos Refúgios indica que o modelo sucumbiu frente ao teste permitido por um banco de dados paleoecológicos, o qual não estava disponível durante a "idade dourada" deste paradigma, quando evidências indiretas eram consideradas satisfatórias para mantê-lo.
Palavras-chave: Amazônia, refúgios, pólen fóssil, glaciação, Pleistoceno, Holoceno, Mioceno, filogenia, diversidade e especiação
Introduction
One of the great biological patterns that ecologists strive to explain is the latitudinal gradient in diversity. The incredible diversity of tropical systems and the tapering of that richness poleward has been observed and debated by Wallace and Darwin and others following in their footsteps. Various hypotheses have been established: that the tropics accumulate species without much extinction (museum hypothesis); that the area of the tropics is greater than at other latitudes (Rosenzweig & Sandlin 1997); that most modern lineages originated in the tropics and relatively few have been able to leave (Ricklefs 1987); and that intermediate levels of disturbance maintain diversity but do not cause extinction (Connell 1978). All of these hypotheses have merit, yet none provides a completely satisfactory answer. What was lacking from all of them was an explanation of why speciation rates may have been higher in a tropical setting than in a temperate one.
Conventional wisdom of the 1960s stated that tropical regions were rich in biodiversity because they were ecologically stable. Climate never changed, therefore species never went extinct and simply accumulated. The stability of tropical climates was challenged by findings in Africa that both precipitation and temperature had changed during the last ice-age (Livingstone 1967). Africa was drier and colder in the last ice age. Forests were forced downslope in the Ruwenzori and grasslands invaded. In the Andes on the High Plain of Bogotá, Colombia, van der Hammen and Gonzalez (1959) described a long pollen history of warm and cold oscillations. Each cold oscillation was an ice age that caused forests to die out at high elevations and to be replaced by Paramo grasslands. These findings of changeable tropical climates and forests being replaced by grassland set the scene for new thinking about tropical lowland systems.
In 1969, Jurgen Haffer, a petroleum geologist and ornithologist, introduced an elegant theory of ice-age speciation events based on climate change (Haffer 1969). He observed that modern bird distributions of closely related species (congeners and superspecies) often had ranges that abutted one another but did not overlap (i.e. parapatric). He also noted that there was a predictable pattern in which certain areas held more endemic species than others. From these observations he made the intellectual leap to observe that for these species to have speciated they must have been spatially separated from their parapatric counterparts at some time in the past. Haffer assumed, as would many biologists, that allopatry (reproductive isolation from other gene pools of the same species) was an essential precursor for speciation. He invoked the new climatic data emerging from Africa of a dry glacial period (ca. 100,000-20,000 years ago) to argue that the Amazon Basin was similarly arid, and it was seas of savanna surrounding islands of forest that provided the genetic isolation required for speciation.
Haffer hypothesized that the wettest areas would be hilltops that caught orographic rainfall, and therefore remained wet enough to support evergreen forest. The interstices between the hilltops became too dry to support forests, and savanna grasslands expanded. Those wettest locations where evergreen forest survived, would have formed isolated refuges for all forest wildlife; hence they were termed refugia. Within these refugia, Haffer suggested, populations of birds, insects, and plants were isolated. Cut off from the populations of other forest organisms in other refugia, the fragmented populations underwent speciation. The model was the first comprehensive attempt to provide a mechanism that would lead to unusually high rates of speciation in some tropical areas.
The elegance of the model attracted many other workers to develop similar refugial maps for butterflies, frogs, lizards, and some families of plants (Vanzolini 1970, Haffer 1974, Prance 1982, Haffer 1985, Brown 1987a, Brown 1987b, Haffer 1991, Haffer & Prance 2001) (Figure 1). The refugial hypothesis is included as the definitive history of Amazonian forests in many texts; however, it is important to note that it is not based on any paleoecological data (Colinvaux 1989). The refugial model and associated Amazonian aridity became a paradigm, but one that was founded on hypothesis not empirical data. We will review the assumptions of the model because the most attractive features of the refugial hypothesis were that it was both explanatory and predictive, and hence testable.
Assumptions of the model
The refugial model has been considerably revised since its first formulation. However, at the core of the model are 6 key assumptions that have remained constant.
1) The biogeographic data are sufficient for hypothesis generation.
2) Process of speciation: all speciation must result from spatial allopatry.
3) The spatial pattern of speciation: Speciation occurred in identifiable centers that are independent of major modern landscape barriers.
4) The temporal pattern of speciation: Amazonian genera (especially the ones used to generate the hypothesis) experienced sudden bursts of speciation centered on the ice ages.
5) Changes in precipitation and seasonality: In the interstices between the hilltops the replacement of forest with savanna would require a shift from the "normal" Amazonian precipitation of 2200-3000 mm and 0-3 months dry season, to a system receiving <1500 mm and a 5 month dry season.
6) Unchanging temperature: For the hilltops to remain suitable for the most sensitive of lowland species requires temperatures in the lowland tropics to be about the same as those of the present during the ice ages.
It can be argued that the latest version of the model relaxes Assumptions 3 and 4 (Haffer & Prance 2001), however, to do so invalidates the original worth of the model as a predictive tool (Bush 2005), and so we include them in this analysis.
Assumption 1: The biogeographic data are sufficient for hypothesis generation. Are the spatial and taxonomic patterns upon which the refugial hypothesis is built real? In an analysis of herbarium collections records (Nelson et al. 1990) showed strong congruence between geographic variation in collecting intensity and apparent species diversity. It is likely that botanists are drawn to locations of high diversity and so the implied pattern of diversity may be real. However, some of the sites of highest intensity were close to major points of entry or centers of research, and thus suggest that the collecting effort at those locations may have been disproportionately high. In truth, most areas of the Neotropics are so incompletely surveyed, and taxonomy is still so fluid for complex groups, that maps of diversity and distribution must be treated with some caution.
In a statistical analysis of the overlap of Amazonian bird distributions, instead of finding refugial centers, no pattern was found beyond randomness (Beven et al. 1984). While our knowledge of species composition and full diversity of any given location is imperfect and this could either strengthen or weaken refugial claims, the overall pattern of species richness across Amazonia is unlikely to alter radically as new data emerge. Hence we will accept Assumption 1. In so doing we accept that the major biogeographic regions of Amazonia (Figure 2a) require some explanation.
Assumption 2: Process of speciation. Ernst Mayr was a staunch advocate of both the importance of allopatry in speciation and in the refugial hypothesis (Mayr & O'Hara 1986). While sympatric speciation through polyploidy has been demonstrated in plants, and post-zygotic isolation is probably possible without allopatry, the consensus among biologists is that allopatry is necessary for speciation (Coyne & Orr 2004). As a detailed review of these arguments is beyond the scope of this paper we will accept this assumption, but note that our understanding of genetic isolation may be radically altered as more molecular studies are conducted on seemingly continuous populations (McLachlan & Clark 2005).
Assumption 3: The spatial pattern of speciation. Modern barriers such as edaphic types, while incorporated into later versions of refugial prediction are not seen as sufficient barriers to cause observed biogeographic patterns. Similarly, the large Amazonian rivers and the Andes mountains are not treated as standing constraints to genetic interchange.
Wallace (1852) and later Endler (1982) proposed that rivers were indeed major barriers to dispersal and subsequent studies have yielded mixed results. For some aquatic, amphibian and riparian taxa the rivers are clearly conduits not constraints (Gascon et al. 2000). Similarly the genetic isolation of a forest-floor dwelling antbird by a river several kilometers wide will be much more likely than for a highly motile bird such as a toucan (both these groups were used as examples of refugial species). Equally, spider monkey populations may be separated at the mouth of a major river, but are not greatly influenced by its headwaters (Ayers and Clutton-Brock 1992, Collins & Dubach 2000).
Phylogenetic studies have been conducted on many groups and some important trends emerge: 1) No single pattern of biogeographic origin is common to all taxa (for an excellent faunal review see Moritz et al. 2000); 2) Most phylogenies have a tree with a basal division that separates Central American and Chocó populations from those of Amazonia and the Atlantic forests (Figure 2b); 3) Genotypes from the Guianan highlands and the Atlantic Coastal forest and the dry forests fringing Amazonia need to be included in the analyses (Costa 2003). The importance of the connectivity of eastern Brazilian ecosystems has also been demonstrated through the study of fossil pollen and speleothem data (de Oliveira et al. 1999, Auler et al. 2004). Again we point to the study by Beven et al. (1984) that found the distribution of taxa to be random rather than falling within predictable centers. None of these studies pinpoint refugial locations, but they do bear out the major biogeographic provinces of Amazonia, albeit with major watersheds shifting between provinces according to the phylogeny. Thus Assumption 3 is neither supported by, nor refuted, by the available data.
Assumption 4: That "Amazonian genera (especially the ones used to generate the hypothesis) experienced sudden bursts of speciation centered on the ice ages". This is an area where research is advancing rapidly, data are still sparse, and there are many interesting issues yet to resolve. However, preliminary data show that speciation events among bats, birds, snakes, and mammals (Bates & Zink 1994, Patton et al. 1994, Zink & Slowinski 1995, Bates et al. 1998, Bates 2000, Patton et al. 2000, WWF 2003, Zink et al. 2004, Gosling & Bush 2005) in Amazonia have been a continuous process, without a clear surge of new forms in the Quaternary.
Phylogenetic trees provide a rough hierarchy of separation. According to the phylogeny, basal splits are generally 8-15 million years ago (Knapp & Mallett 2003). The orogeny of the Andes progressively isolated Amazonia from the Chocó and Central America on a similar timeframe (Hoorn et al. 1995). The rise of the Andes not only caused vicariance among populations, but also changed the drainage of Amazonia causing the formation of the modern mighty river system. This period of 20 to10 million years ago was also a time of a marine highstand with sea-level probably at its highest around 14 million years ago (30-50 m above present level). That highstandcoupled with forebasin downwarping, resulted in extensive epicontinental seas within Amazonia (Figure 3). While some debate exists regarding the full extent and connectivity of these water bodies (e.g. Hoorn et al. 1995, Rasanen et al. 1995), it is apparent that they would have been potentially much greater barriers to dispersal than any modern river of the Amazon Basin (Rasanen et al. 1995, Hovikoski et al. 2005).
Much of the proto Andes would have supported lowland forests prior to the mid-Miocene and these areas may subsequently have provided species for the expanding western Amazon forests that colonized land left by falling mid-Miocene to late-Miocene sea-levels.
Other major changes took place as epicontinental seas in eastern and southern Amazonia formed and subsequently drained (Nores 1999, Hovikoski et al. 2005). The upwarp of the Andes and the drainage of the Pebasian Sea were closely linked. Lineages previously to the west of the Pebasian and Paranense Seas, and separate from eastern Amazonia, might account for the basic east-west biogeographic split in some Amazonian clades. Similarly the seaway or the great wetlands if this were not an actual sea, that lay in the modern Amazon channel could have provided the basic north-south discontinuity. As we realize that many modern clades are rooted in the Miocene it is important to build these ancient landscapes into our evolutionary vision (Figure 3).
The phylogenetic data clearly reveal that there was no sudden wave of glacially-induced speciation in Amazonia and that focusing on the Quaternary as a source of Amazonian diversity is temporally myopic. This realization clearly violates Assumption 4 of the refugial hypothesis.
Assumptions 5 and 6: The past ice-age climates of Amazonia. A discussion of these data follows, but it is not quite so straightforward as those given above, hence the data will be presented in a somewhat different format in which the view of the original author is reported and then we provide an updated re-interpretation or commentary.
Paleoecological data
In a perfect world, the refugial hypothesis would be simple to test. Lake-sediment core samples would yield fossil pollen that would allow description of regional vegetation through time. Replicated core samples that spanned the last ice age would be collected from areas inside and outside of postulated refugia. If the refugial samples were not always lowland forest, or if the outside samples did not show an oscillation to grassland during ice ages, then the hypothesis would be falsified.
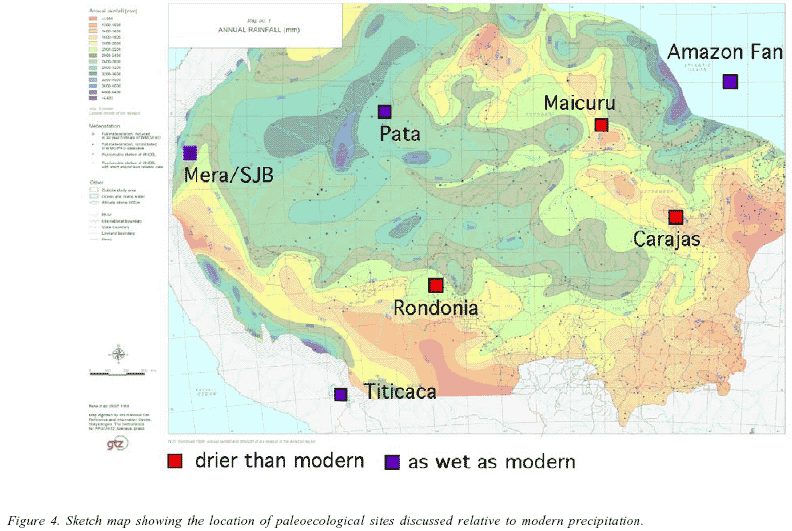
Despite 30 years of actively searching for such sites, paleoecologists still have not found enough ancient lakes in appropriate settings to make this a simple test of Assumptions 5 and 6. The processes that provide ancient lakes in temperate areas, such as glacial activity, solution basins, and volcanoes are largely absent from the Amazon Basin. Furthermore vigorous bankside erosion by vast meandering rivers, obliterates lakes in floodplains every few thousand years. Only a handful of lakes of sufficient age to test the refugial hypothesis have been found in Amazonia. These lakes are all somewhat ecotonal relative to proposed refugia (Figure 4). The fossil records of these systems provide detailed histories of those sites, but they have to be interpreted in a larger context. In other words, just because you see a pattern consistent with, or contrary to, the expectations of the refugial hypothesis at a single marginal location, it does not substantiate or disprove the hypothesis. Ecotonal boundaries can migrate 10s even a 100 kilometers or more without indicating that the 1000's of kilometers of forest were similarly impacted.
We briefly review sites that provide the principal evidence for and against refugia on the basis of paleoecology and distinguish the author's interpretation from our commentary on the record.
Rondônia, Brazil
This area currently supports tropical rain forest, but the refugial hypothesis predicts that it would be savanna during ice age times. A fossil pollen record (van der Hammen 1974) that shows an oscillation between modern forest and grassland (Figure 5). Proportions of Poaceae (grass) pollen in this sequence are consistent with those documented from modern savanna habitats. Undoubtedly, this is the best palynological evidence to support the existence of refugia. The forest is clearly Holocene in age, while the grassland episode is undated but assumed to be part of a continuous depositional sequence and therefore attributable to the terminal portion of the ice age. The location of the site lies outside of any proposed refugium, and so this meets prediction 3 of the refugial hypothesis.
Our thoughts on the Rondônia sequence:
There is little doubt that this site once supported a grassland, but there is no datum to suggest when this occurred. Below, we will discuss the frequency of climate change in the Amazon Basin, and demonstrate that wet, dry, cold and warm climatic oscillations have occurred with great rapidity. Northern Amazonia is shown to have had active dune fields in some areas with a lot of activity between 17,000 BP and 8000 BP (Filho et al. 2002). This dune activity implies reduced precipitation, between 17,000 and 8,000 BP. The Rondônia site lies within 100 km of the modern savanna-forest ecotone and has a precipitation pattern that is only just sufficient to support closed canopy forest. In a location as close to an ecotone as this site, the drought at 15,000 BP could easily account for the undated expansion of savanna seen in this record.
What cannot be concluded from this site is that a) the transition to savanna lasted an evolutionarily significant amount of time; and b) that other, less ecotonal regions, were similarly affected.
Carajas, Brazil
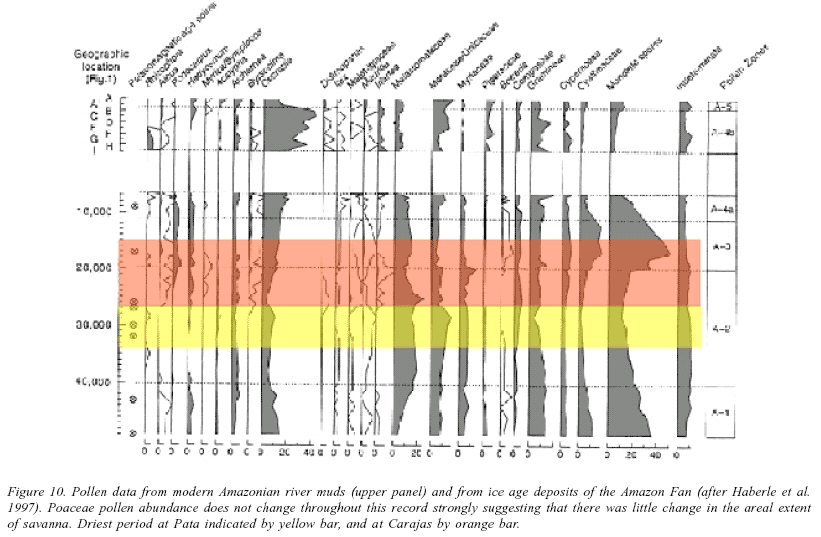
The Carajas record comes from a 600 m high inselberg that rises above the Amazonian plain in south-eastern Amazonia that presently supports a mixture of savanna and open woodland habitats. This area was predicted to lie outside of any ice-age refugium.
The fossil pollen and paleolimnological data from this site exhibit a dry interval between ca. 26,000 and 15,000 BP (Absy et al. 1991, Sifeddine et al. 2001). In this episode the lake dried out, and the last pollen signature before a total gap in sedimentation is rich in Poaceae (grass), Asteraceae, and Borreria pollen (Figure 6). These three pollen types are used as indicators of the presence of savanna. Consequently, these data were interpreted to indicate a widespread savanna expansion at the LGM, fully consistent with the refugial hypothesis (Absy et al. 1991, Haffer & Prance 2001).
Our thoughts on the Carajas sequence:
An alternate interpretation of this data set accepts that there was a dry period between 26,000 and 15,000 BP, but questions whether the Asteraceae, Borreria, and Poaceae indicate savanna conditions. As a lake contracts it offers a smaller and smaller surface area onto which pollen falls. It is widely accepted that the smaller the lake surface area, the stronger is the input of local pollen types. In other words, large lakes (>100 m in diameter provide regional pollen records, whereas small lakes <20 m provide an image of the immediately adjacent marsh and little else (Jacobson and Bradshaw 1981, Prentice 1985). Thus, as the lake at Carajas dried out, the marsh plants (Poaceae, Asteraceae, and Borreria) increased in proportion in the pollen record, without necessarily influencing the local forest. Do we believe that you can dry out a lake on an inselberg that presently supports woody savanna and not have an expansion of grass savanna? Probably not. It is extremely likely that on that dry hilltop there was local expansion of savanna, but this does not tell us anything about what was happening in the wetter lowlands. Note also that the highest percentages of Poaceae pollen do not occur at the LGM but during the Holocene. No-one has suggested the existence of savanna in Holocene times, and so it appears that Poaceae pollen abundance is not directly correlated with the presence of refugia (Bush 2002).
A further point to consider is what if we are wrong? What if the drying really did last from 26,000 BP to 15,000 BP and substantial areas of savanna spread into Amazonia, does this provide time enough for allopatric speciation? Turning this argument around we can say that a roughly equivalent time has passed in the Holocene (11,000 years) and ask if there has been a radiation of isolated savanna species within the Amazon Basin (there are large modern savanna islands within a sea of forests). The answer is, there has been no such speciation. Very rapid speciation has been reported for a few organisms such as African cichlids and fruit flies, but neither of these are well-defined, and where there is very tight co-evolution (Coyne & Orr 2004). In summary, it is very unlikely that 11,000 years of isolation is enough to cause widespread speciation among butterflies and birds, let alone among such long-lived organisms as trees. The evidence for drying in ice-age Amazonia indicates neither the intensity nor the duration sufficient to bring about allopatric speciation.
The Carajas data, rather than supporting the refugial hypothesis, are in fact a further refutation of Prediction 5 that Amazonian genera (especially the ones used to generate the hypothesis) experienced sudden bursts of speciation centered on the ice ages.
In a more recent paper, additional information was released about the Carajas record (Ledru et al. 2001) indicating that a significant amount of Podocarpus pollen was also found in the ice-age sediments. Podocarpus is a gymnosperm tree that is most abundant in cloud forest above 1500-2200 m elevation. For Podocarpus to occur at this site strongly suggests a cooling at the LGM. In the tropics a rule of thumb is that a 1000 m increase in altitude results in a 5ºC drop in temperature. For Podocarpus to be found about 1000 m below its normal range suggests about a 5ºC cooling at the LGM. This cooling (originally denied for this site) is now seen as consistent with many other regional records (Colinvaux 1987, Bush et al. 2001). A 5ºC cooling may not sound all that much, but it is the equivalent of trading the climate of Atlanta with that of Washington DC, or Berlin with that of Moscow.
In summary, the data from Carajas refute Assumption 5 and the observation that there was regional cooling refutes Assumption 6 that ice-age temperatures were similar to those of the present.
Paleoecological data published that opposed refugia
Mera and San Juan Bosco, Ecuador
These two sites lie within the area of the postulated Napo refugium. For the refugial hypothesis to be true they should present an unchanging history of lowland forests.
However, these sites provided the first direct evidence that the lowlands were moist and cool during the latter part of the ice age. Now, better records exist for other locations, but these sites are important as they represented the turning of the tide.
Mera (Liu & Colinvaux 1985) and San Juan Bosco (Bush et al. 1990) lie at the foot of the Andes, at 1100m and 970 m respectively, immediately above the great Amazonian plains. These records are both cliff exposures in which a downcutting river has exposed ancient sediments. Podocarpus trunks poked out of the cliff and these provided the basis for secure radiocarbon ages spanning ca. 38,000 to 30,000 cal. yr BP (33,000 to 26,000 14C BP). The sediments around the wood were fine silts, indicating a very low energy depositional system (lake or marsh). Pollen and macrofossils of Podocarpus, Drimys, Alnus (alder), and other montane taxa were very abundant, particularly in the older sediments (Figure 7).
Grass phytoliths (silica bodies inside leaves of grasses) that were present were from C3 grasses not from C4 grasses. None of these plants currently grow below 1800-2500 m elevation in this section of the Andes, and all are found in moist cloud forest environments. These data strongly suggest a 5 ºC cooling and abundant moisture at the foot of the Andes in the last ice age.
The Hill of Six Lakes Brazil
This 300 m high hill rises out of the northern Amazon plain and lies close to the boundary of one of the proposed refugia. The soils on the hill are very thin resulting in an edaphically dry woodland. Some dispute exists regarding whether this site is inside or outside of proposed refugia (Colinvaux et al. 2001, Haffer & Prance 2001). The boundaries of refugia have changed as maps are redrawn and in their latest manifestation Haffer and Prance state that this Hill lies within a refugium. However, the criteria to determine refugia are based on the overlap of endemic species, precipitation and soils. The thin soils of this hill result in a rather species poor, small stature forest, for which there is no evidence of high proportions of endemics. We will treat this as an ecotonal setting for which there are no expectations.
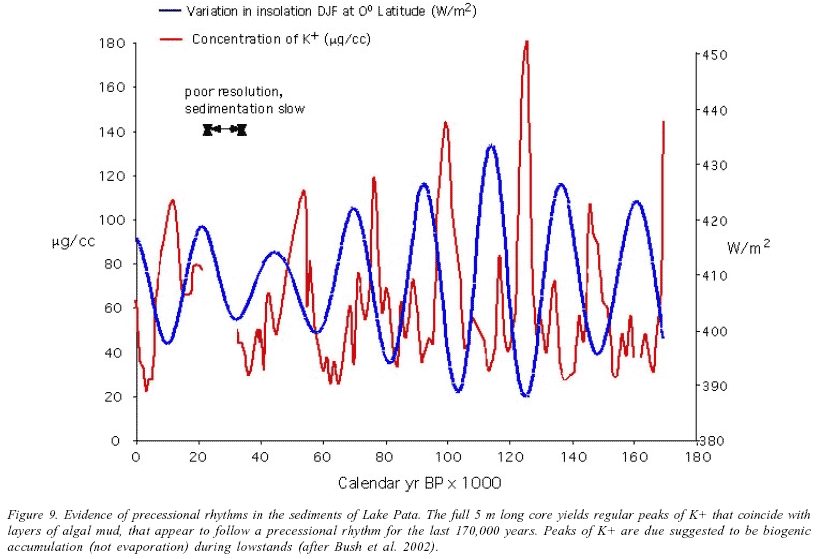
Three lake records have been analyzed, and all show similar histories. The pollen diagram from Lake Pata is representative and it shows the continuous presence of forest on this site throughout the last 50,000 years (Colinvaux et al. 1996). However, the forest did not remain unaltered and, as at Carajas, Mera and San Juan Bosco, the presence of cold elements Podocarpus, Hedyosmum, Weinmannia, Myrsine, and Ericaceae, strongly suggests a 5ºC cooling (Figure 8). Pata also revealed an intriguing pattern of lake-level changes, which allow us to draw inferences regarding changes in precipitation. Lake Pata is a small, shallow (mostly 3m deep) closed basin lake. Based on sedimentary oxidation, exceptionally high pollen concentrations, and algal blooms, a series of low-lake stands are evident in the Pata record (Bush et al. 2002). During each of these events there is a peak of K+ cations in the sediment. Potassium normally weathers out of local rock at the same rate as sodium, and so the two concentrations normally co-vary. However, at Pata there appear a rhythmic set of K+ peaks that are independent of Na+ concentrations. The most probable explanation is that during times of low lake level the photic zone of the lake extended down to the lake bed. Under these circumstances algae can access nutrients in the lake mud, and the system switches from being oligotrophic to eutrophic. Algae stored the K+, but would not store Na+, and so when the algal blooms die off there is increased concentration of K+ in the mud. If this process is repeated for several thousand years there develops a significant spike of K+ concentration.
A robust chronology based on 15 radiocarbon AMS dates shows that the driest time at Pata was between 35,000 and 23,000 BP. At this time the lake was so reduced that repeated sedimentary oxidation prevented net accrual of sediment. If the basal age of the core is calculated on extrapolation of the sedimentation rate in the AMS dated portion of the core, a basal age of 180,000 yr BP is suggested. Through the last 40,000 years the peaks of K+ coincide with the dry season, June-July-August (JJA), insolation maxima. If the peaks of K+ are assumed to follow the pattern observed in the radiocarbon dated section of the core, orbitally tuning the remaining peaks to JJA insolation maxima provides a basal age of 170,000 BP (Figure 9). The K+ peaks are lowstand events and the intervening periods of low K+ concentrations are wetter periods that coincide with the December-January-February insolation maxima.
To generate Figure 9, the only tuning beyond the range of the 14C record was to align the seventh K+ peak with the seventh insolation peak, all other samples were allowed to fall without adjustment, i.e. an even rate of sedimentation is assumed between 45,000 and 170,000 years. It is evident from this record that the cyclic droughts show a remarkable concurrence with the JJA insolation and of a weaker set of dry events coinciding with the DJF curve. This pattern faithfully replicates the observed relationship between seasonal insolation and lake level on the Bolivian Altiplano (Baker et al. 2001, Fritz et al. 2004, Chepstow-Lusty et al. 2005).
In the dry events at Lake Pata we know that lake level dropped, indicating a decrease in precipitation, yet the forest was not replaced by grassland. We hypothesize that the principal reduction in precipitation was in the wet season. During the wet season there is excess water in this system, some of which enters the lake and raises lake level. However it is the dry season and the thin soils that determine vegetation type. If wet season rains were reduced, it is probable that there would be little change in forest composition, but lake level would respond. Had it been a dry season reduction in precipitation the forest would have converted to grassland, and the system would have become fire-prone but there is no cyclic occurrence of charcoal or reduction in forest cover to suggest such a pattern.
The paleoecological records from two other lakes, Verde and Dragao, on the Hill of Six lakes have now been published (Bush et al. 2004) and these are entirely consistent with the history from Lake Pata, though neither provides as sensitive a cation record. From these additional records the peak of the late glacial dry event is confirmed to have occurred between 35,000 and 23,000 cal. yr BP. The analysis of thee lakes provided an additional insight into the low-lake stands observed at Pata.
Lake Dragao was (and is) highly susceptible to lake level change. In a two-week field operation we observed its water level go down by 2 m much more than could be explained by evaporation alone. A geological team played soccer on its dried out lake bed in the 1982 El Nino drought. Clearly this lake is leaking and its hydrology is a finely tuned balance between input (rainfall) and leakage. A clear inference is that lakes more susceptible to drought will contain greater durations of sedimentary hiatus in their history. Indeed Dragao has an apparent hiatus until c. 18,640 cal year BP, Similarly, Lake Verde, which is a nine meter deep lake today failed to accumulate sediment throughout the Holocene. If this lack of sedimentation is genuine and not an artefact of coring, it is probable that Verde has only recently refilled with water in the last millennium or so. Being dry in the early and mid Holocene, would be predicted from the precessional pattern. However, there is no suggestion of forest loss in the Holocene.
Again lake-level on these inselberg lakes is shown to be a sensitive proxy for a net change in the precipitation-evaporation-leakage balance, but an unreliable proxy for inferring vegetation change.
In summary: The lakes of the Hill of Six Lakes provide evidence of cooling, and establish that precipitation patterns oscillated cyclically, did not change in the manner predicted by the refugial hypothesis.
Maicuru
Maicuru (Colinvaux et al. 2001) is an inselberg that lies at 0º latitude in eastern Amazonia. This Hill rises to 500 m and has numerous small, shallow, lakes on its summit plateau. The largest of these lakes provided another long but discontinuous paleoecological record that indicates the presence of forest throughout the represented portion of the last ice age. In this record the peak of the last ice age is missing, as there is a sedimentary hiatus between 30,000 and 15,000 BP. Thus this record is palynologically similar to that of the Hill of Six lakes, but the duration of the dry event looks to be somewhat longer, starting at about the same time as the Hill of Six Lakes and ending at about the same time as Carajas, making it a longer climatic feature.
Data from the Amazon Fan
The Amazon river acts as a vast pollen trap, collecting pollen from the entire Amazon Basin. Pollen is carried seaward in the river water. So that at the Atlantic coast, the pollen of the turbid Amazon waters represents the vegetation types of the entire subcontinental area of Amazonia. When the riverwater discharges into the ocean, the sudden reduction in flow rate causes the sediments and pollen to be deposited. Sediment cores from the Amazon fan provide an insight on the past vegetation of Amazonia at the scale of that landscape. Simon Haberle undertook a study of modern sediments in Amazonian tributaries (upper panel Figure 10) and of fossil sediments raised from the Amazon fan (Haberle 1997).
These data show that in the modern rivers Poaceae pollen accounts for about 10% of the pollen sum, and that the ice age pollen spectra were similarly low in Poaceae pollen.
These data clearly demonstrate the fallacy that large areas of savanna replaced forest.
Haffer & Prance (2001) and van der Hammen & Hoogiemstra (2000) argue that the Amazon fan data only represent riparian vegetation and that there is no information in this data set about the Amazon Basin as a whole. The problem with this interpretation is that the Amazon and its major tributaries are huge rivers, several kilometers in width, which will trap pollen at the regional scale not the local scale. The source of pollen will be water draining from a vast riveraine network, and also airborne pollen that is scrubbed from the atmosphere during the frequent convective storms. To argue that this pollen record essentially riparian as opposed to regional is most improbable.
Conclusions
The Pata record shows very clearly that tropical climate change is not a simple switch between glacial and interglacial conditions. From studies of fossil pollen and plant remains in South and Central America, Africa, and Asia, it has become clear that ice ages cannot simply be classified as warm or cold, wet or dry. Ice ages were times of overall cooling in which there occurred warm, dry and wet events that lasted decades to millennia. Simple models portraying any kind of uniform condition will be wrong.
Furthermore, in an area as vast as Amazonia climate change was geographically heterogeneous, so that not all areas would have been experiencing drought, or flood, at the same time. For example, the low lake level event at Pata was between 35,000 and 26,000 BP whereas the low lake event at Carajas was between 26,000 and 15,000 BP.
And then there is the Maicuru dry event that appears to be a combination of both Pata and Carajas, however, whether the Maicuru drying is genuinely different from that of the other lakes, or if it is simply a function of a shallower lake in a drier part of Amazonia, remains to be tested. What is evident from the Hill of Six lakes is that though the three records show subtly different lengths of the driest of late Pleistocene events, the forest is not necessarily greatly altered. For example, the onset of the dry event at Verde is beset by several reversals in the 14C record and cannot be reliably dated after about 40,000 cal. yr BP. However, despite this obviously being a period of low lake level at Verde, the records from Pata and Dragao (within 5 km of Verde) show no significant change in the forest. Thus simply because lake level falls does not mean that forest disappears.
The timing of inferred lake-level at the Hill of Six Lakes fits very well with regional changes in convection (Bush & Silman 2004) and meshes well with speleothem data from Rio de Janeiro (Cruz et al. 2005), and lake level in the High Andes (Baker et al. 2001). In both these extra-Amazonian records climate is driven by wet-season (December-February) precessional forcing, exactly the periodicity of highstands and lowstands observed at Pata (Bush 2005).
In conclusion, we observe that the refugial hypothesis fails on Assumptions 4-6 and is not supported by available molecular or paleoecological data. Indeed, the latest manifestation of the refugial hypothesis, which relaxes Assumptions 3 and 4, and further modifies the matrix separating refugia from savanna to dry forest or riparian corridors (Haffer & Prance 2001), provides no mechanism to engender widespread allopatry and speciation.
If after almost 40 years we refute the refugial hypothesis as an explanation of Amazonian diversity, have we come full circle, and arrived back at the starting position of 1969? We do not believe that is the case. In the last 30 years the scientific community has done exactly what good scientists do, we have tested a good initial hypothesis by gathering a vast amount of data, and then rejected the hypothesis. We know much more about the history of Amazonia, the way that Amazonian climate works, and about the evolutionary history of a broad range of species, than at the start of this quest. We have rejected a simple hypothesis of speciation for a more realistic understanding of the complexity of evolutionary and climatic processes. We understand more about the migration of species, and the transience of tropical plant communities and can apply this new knowledge to the pressing issues of global climate change and conservation biology. But there is so much more to know!
Acknowledgements
This work was supported by National Science Foundation Grants BSR 9007019 and DEB9732951. We are indebted to our long-term collaborator Paul Colinvaux for his support, encouragement and insights. Very sincerely we wish to thank Jurgen Haffer for his visionary hypothesis that has promoted so much research.
Date Received 05/12/2005
Revised 10/20/2005
Accepted 01/01/2006
ISSN 1676-0611
- ABSY, M.L., CLIEF, A., FOURNIER, M., MARTIN, L., SERVANT, M., SIFEDDINE, A., SILVA, F.D., SOUBIÈS, F., SUGUIO, K.T. & VAN DER HAMMEN, T. 1991. Mise en évidence de Quatre phases d'ouverture de la forêt dense dans le sud-est de L'Amazonie au cours des 60,000 dernières années. Première comparaison avec d'autres régions tropicales. Comptes Rendus Academie des Sciences Paris, Series II 312:673-678.
- AULER, A. S., WANG, X., EDWARDS, R.L., CHENG, H., CRISTALLI, P.S., SMART, P.L., & RICHARDS, D.A. 2004. Palaeoenvironments in semi-arid northeastern Brazil inferred from high precision mass spectrometric speleothem and travertine ages and the dynamics of South American rainforests. Speleogenesis and Evolution of Karst Aquifers 2:1-4.
- AYERS, J. M., & CLUTTON-BROCK, T.H.. 1992. River boundaries and species range size in Amazonian primates. Am. Nat. 140:531-537.
- BAKER, P. A., RIGSBY, C.A., SELTZER, G.O., FRITZ, C.S., LOWENSTEIN, T.K., BACHER, N.P., & VELIZ, C. 2001. Tropical climate changes at millennial and orbital timescales on the Bolivian Altiplano. Nature 409:698-701.
- BATES, J. M. 2000. Allozymic genetic structure and natural habitat fragmentation: Data for five species of Amazonian forest birds. Condor 102:770-783.
- BATES, J. M., HACKETT, S.J., & CRACRAFT, J. 1998. Area-relationships in the Neotropical lowlands: an hypothesis based on raw distributions of Passerine birds. J. Biogeogr. 25:783-794.
- BATES, J. M., & ZINK, R.M. 1994. Evolution into the Andes: molecular evidence for species relationships in the genus Leptopogon Auk 111:507-515.
- BEVEN, S., CONNER, E.F., & BEVEN, K. 1984. Avian biogeography in the Amazon Basin and the biological model of diversification. J. Biogeogr. 11:383-399.
- BROWN, K.S.J. 1987a. Conclusions, synthesis and alternative hypotheses. In Biogeography and Quaternary history in tropical America (T.C. Whitmore & G.T. Prance, eds). Oxford Science Publications, Oxford, p.175-196.
- BROWN, K.S.R., JR. 1987b. Biogeography and Evolution of Neotropical Butterflies. In Biogeography and Quaternary history in tropical America (T.C. Whitmore & G.T. Prance, eds). Oxford Science Publications, Oxford, p.66-104.
- BUSH, M. B. 2002. On the interpretation of fossil Poaceae pollen in the humid lowland neotropics. Palaeogeogr. Palaeocl. 177:5-17.
- BUSH, M. B. 2005. Of orogeny, precipitation, precession and parrots. J. Biogeogr. 32:1301-1302.
- BUSH, M.B., MILLER, M.C., DE OLIVEIRA, P.E. & COLINVAUX, P.A. 2002. Orbital forcing signal in sediments of two Amazonian lakes. Journal of Paleolimnology 27:341-352.
- BUSH, M.B. & SILMAN, M.R. 2004. Observations on Late Pleistocene cooling and precipitation in the lowland Neotropics. J. Quaternary Sci. 19:677-684.
- BUSH, M.B., DE OLIVEIRA, P.E., COLINVAUX, P.A., MILLER, M.C. & MORENO, E. 2004. Amazonian paleoecological histories: one hill, three watersheds. Palaeogeogr. Palaeocl.
- BUSH, M.B., STUTE, M., LEDRU, M.-P., BEHLING, H., COLINVAUX, P.A., DE OLIVEIRA, P.E., GRIMM, E.C., HOOGHIEMSTRA, H., HABERLE, S., EYDEN, B.W., SALGADO-LABOURIAU, M.-L. & WEBB, R.. 2001. Paleotemperature estimates for the lowland Americas between 30şS and 30şN at the last glacial maximum. In Interhemispheric Climate Linkages: Present and Past Interhemispheric Climate Linkages in the Americas and their Societal Effects (V. Markgraf, ed.). Academic Press, New York. p.293-306.
- BUSH, M.B., WEIMANN, M., PIPERNO, D.R., LIU, K.-B. & COLINVAUX, P.A. 1990. Pleistocene temperature depression and vegetation change in Ecuadorian Amazonia. Quaternary Res. 34:330-345.
- CHEPSTOW-LUSTY, A.J., BUSH, M.B., FROGLEY, M.R., BAKER, P.A., FRITZ, S.C. & ARONSON, J. 2005. Vegetation and climate change on the Bolivian Altiplano between 108,000 and 18,000 yr ago. Quaternary Res. 63:90-98.
- COLINVAUX, P.A. 1987. Amazon diversity in the light of the paleoecological record. Quaternary Science Rev 6:93-114.
- COLINVAUX, P.A. 1989. The past and future Amazon. Scientific American:101-108.
- COLINVAUX, P.A., DE OLIVEIRA, P.E., MORENO, J.E., MILLER, M.C. & BUSH, M.B.. 1996. A long pollen record from lowland Amazonia: Forest and cooling in glacial times. Science 274:85-88.
- COLINVAUX, P.A., IRION.G., RÄSÄNEN, M.E., BUSH, M.B. & NUNES DE MELLO, J.A.S. 2001. A paradigm to be discarded: geological and paleoecological data falsify the Haffer and Prance refuge hypothesis of Amazonian speciation. Amazoniana 16:609-646.
- COLLINS, A. C., AND J. M. DUBACH. 2000. Biogeographic and Ecological Forces Responsible for Speciation in Ateles International J. Primatol. 21:421-444.
- CONNELL, J. H. 1978. Diversity in tropical rain forests and coral reefs. Science 199:1302-1310.
- COSTA, L.P. 2003. The historical bridge between the Amazon and the Atlantic Forest of Brazil: a study of molecular phylogeography with small mammals. J Biogeogr. 30:71-86.
- COYNE, J.A. & ORR, H.A.. 2004. Speciation. Sinauer Associates, Sunderland, Massachussetts.
- CRUZ, F.W., JR, BURNS, S.J., KARMANN, I., SHARP, W.D., VUILLE, M., CARDOSO, A.O., FERRARI, J.A., SILVA DIAS, P.L. & VLANA,O. JR. 2005. Insolation-driven changes in atmospheric circulation over the past 116,000 years in subtropical Brazil. Nature 434:63-66.
- DE OLIVEIRA, P.E., BARRETO, A.M.F. & SUGUIO, K. 1999. Late Pleistocene/Holocene Climatic and Vegetational History of the Brazilian Caatinga: the fossil dunes of the middle São Francisco River. Palaeogeogr. Palaeocl. 152:319-337.
- ENDLER, J.A. 1982. Pleistocene forest refuges: fact or fancy? Pages 641-657 in G. T. Prance, editor. Biological diversification in the tropics. Columbia University Press, New York.
- FILHO, A.C., SCHWARTZ, D., TATUMI, S.H. & ROSIQUE, T. 2002. Amazonian paleodunes provide evidence for drier climate phases during the late Pleistocene-Holocene. Quaternary Res. 58:205-209.
- FRITZ, S.C., BAKER, P.A., LOWENSTEIN, T.K., SELTZER, G.O., RIGSBY, G.A., DWYER, G.S., TAPIA, P.M., ARNOLD, K.K., KU, T.L. & LUO, S. 2004. Hydrologic variation during the last 170,000 years in the southern hemisphere tropics of South America. Quaternary Res. 61:95-104.
- GASCON, C., MALCOLM, J.R., PATTON, J.L., DA SILVA, M.N.F., BOGART, J.P., LOUGHEED, S.C., PERES, C.A., NECKEL, S. & BOAG, P.T.. 2000. Riverine barriers and the geographic distribution of Amazonian species. P. Natl. Acad. Sci. USA 97:13672-13677.
- GOSLING, W.D. & BUSH, M.B. 2005. A biogeographic comment on: Wüster et al. (2005) Tracing an invasion: landbridges, refugia, and the phylogeography of the Neotropical rattlesnake (Serpentes: Viperidae: Crotalus durissus). J. Mol. Ecol. 14:3615-3617.
- HABERLE, S.G. 1997. Late Quaternary vegetation and climate history of the Amazon Basin: correlating marine and terrestrial pollen records. In Proceedings of the Ocean Drilling Program, scientific results (R.D. Flood, D.J.W. Piper, A. Klaus & L.C. Peterson, eds). Ocean Drilling Program, College Station, Texas. p.381-396.
- HAFFER, J. 1969. Speciation in Amazonian forest birds. Science 165:131-137.
- HAFFER, J. 1974. Avian Speciation in Tropical South America. Nuttall Ornithological Club, Harvard University, Cambridge.
- HAFFER, J. 1985. Avian Zoogeography of the Neotropical lowlands. Ornithological Monographs 36:113-146.
- HAFFER, J. 1991. Avian species richness in tropical South America. Studies in Neotropical Fauna and Environment 25:157-183.
- HAFFER, J. & PRANCE, G.T. 2001. Climatic forcing of evolution in Amazonia during the Cenozoic: On the refuge theory of biotic differentiation. Amazoniana 16:579-608.
- HOORN, C., GUERRERO, J., SARMIENTO, G.A. & LORENTE, M.A. 1995. Andean tectonics as a cause for changing drainage patterns in Miocene northern South America. Geology 23:237-240.
- HOVIKOSKI, J., RÄSÄNEN, M., GINGRAS, M., RODDAZ, M., BRUSSET, S., HERMOZA, W., ROMERO PITTMAN, L. & LERTOLA, K. 2005. Miocene semidiurnal tidal rhythmites in Madre de Dios, Peru. Geology 33:177-180.
- JACOBSON, G.L. & BRADSHAW, R.H.W. 1981. The selection of sites for paleovegetational studies. Quaternary Res. 16:80-96.
- KNAPP, S. & MALLETT, J. 2003. Refuting refugia? Science 300:71-72.
- LEDRU, M.P., CORDEIRO, R.C., LANDIM, J.M., MARTIN, L., MOURGUIART, P., SIFEDDINE, A. & TURCQ, B. 2001. Late-glacial cooling in Amazonia inferred from pollen at Lagoa do Caçó, Northern Brazil. Quaternary Res. 55:47-56.
- LIU, K. & COLINVAUX, P.A. 1985. Forest changes in the Amazon Basin during the last glacial maximum. Nature 318:556-557.
- LIVINGSTONE, D.A. 1967. Postglacial vegetation of the Ruwenzori Mountains in equatorial Africa. Ecol. Monogr. 37:25-52.
- MAYR, E. & O'HARA, R.J. 1986. The biogeographic evidence supporting the Pleistocene forest refuge hypothesis. Evolution 40:55-67.
- MCLACHLAN, J.S. & CLARK, J.S. 2005. Molecular indicators of tree migration capacity under rapid climate change. Ecology 86:2088-2098.
- MORITZ, C., PATTON, J.L., SCHNEIDER, C.J. & SMITH, T,B. 2000. Diversification of rainforest faunas: An integrated molecular approach. Annu. Rev. Ecol. Syst. 31:533-563.
- NELSON, B.W., FERREIRA, C.A.C., DA SILVA, M.F. & KAWASAKI, M.L.. 1990. Endemism centers, refugia and botanical collection density in Brazilian Amazonia. Nature 345:714-716.
- NORES, M. 1999. An alternative hypothesis for the origin of Amazonian bird diversity. J. Biogeogr. 26:475-485.
- PATTON, J.L., DA SILVA, M.N.F. & MALCOLM, J.R. 1994. Gene genealogy and differentiation among arboreal spiny rats (Rodentia: Echymidae) of the Amazon Basin: a test of the riverine barrier hypothesis. Evolution 48:1314-1323.
- PATTON, J.L., DA SILVA, M.N.F. & MALCOLM, J.R. 2000. Mammals of the Rio Juruá and the evolutionary and ecological diversification of Amazonia. Bulletin of the American Museum of Natural History 244.
- PRANCE, G.T. 1982. Biological Diversity in the Tropics. Columbia University Press, New York.
- PRENTICE, I.C. 1985. Pollen representation, source area, and basin size: Toward a unified theory of pollen analysis. Quaternary Res. 23: 76-86.
- RÄSÄNEN, M.E., LINNA, A.M., SANTOS, J.C.R. & NEGRI, F.R. 1995. Late Miocene tidal deposits in the Amazon foreland basin. Science 269:386-390.
- RICKLEFS, R.E. 1987. Community Diversity: Relative Roles of Local and Regional Processes. Science 235:167-171.
- ROSENZWEIG, M.L. & SANDLIN, E.A.. 1997. Species diversity and latitudes: listening to area's signal. Oikos 80:172-176.
- SIFEDDINE, A., MARTIN, L., TURCQ, B., VOLKMERRIBEIRO, C., SOUBIES, F., CORDEIRO, R.C. & SUGUIO, K.. 2001. Variations of the Amazonian rainforest environment: A sedimentological record covering 30,000 years. Palaeogeogr. Palaeocl. 168:221-235.
- VAN DER HAMMEN, T. 1974. The Pleistocene changes of vegetation and climate in tropical South America. J Biogeogr 1:3-26.
- VAN DER HAMMEN, T. & GONZÁLEZ, E. 1959. Historia de clima y vegetación del Pleistocene Superior y del Holoceno de la Sabana de Bogotá y alrededeores. Dept. Cundinamarca. Official Report of the Servicio Geologico Nacional, Bogotá.
- VANZOLINI, P.E. 1970. Zoologia sistematica, geografia e a origem das especies. Instituto Geografico São Paulo. Serie Teses e Monografias 3:1-56.
- WALLACE, A.R. 1852. On the monkeys of the Amazon. P Zool. Soc. Lon. 20:107-110.
- WWF. 2003. Ecoregions. in World Wildlife Fund.
- ZINK, R.M., KLICKA, J. & BARBER, B.R. 2004. The tempo of avian diversification during the Quaternary. Philosophiocal Transactions of the Royal Society, London. ser. B 359:215-220.
- ZINK, R.M. & SLOWINSKI, J.B. 1995. Evidence from molecular systematics for decreased avian diversification in the Pleistocene Epoch. P. Natl. Acad. Sci. USA 92:5832-5835.

Publication Dates
-
Publication in this collection
16 Oct 2006 -
Date of issue
2006
History
-
Accepted
01 Jan 2006 -
Received
05 Dec 2005 -
Reviewed
20 Oct 2005