Abstracts
A new species of Hypostomus Lacépède is described from the rio Paraguaçu basin, Bahia State, Brazil. The new species is distinguished from its congeners by having black and conspicuous dots on a pale background, which are similar in size on the head, trunk, and fins, along with ventral surface of head and abdomen naked or the latter plated exclusively on its anterior portion, absence of ridges on head and trunk, and caudal-fin lobes relatively similar in length. The new species further differs from the sympatric H. chrysostiktos by having seven branched dorsal-fin rays instead of 10-11 and represents the eleventh siluriform species endemic to the rio Paraguaçu basin.
Coastal drainages; Hypostominae; Hypostomus chrysostiktos; Northeastern Brazil; Taxonomy
Uma espécie nova de Hypostomus Lacépède é descrita da bacia do rio Paraguaçu, estado da Bahia, Brasil. A espécie nova distingue-se de suas congêneres por apresentar corpo com manchas pretas e conspícuas sobre fundo claro de tamanho semelhante no tronco, cabeça e nadadeiras, além da superfície ventral da cabeça e abdômen nus ou apenas porção anterior do abdômen coberto por placas, ausência de quilhas na cabeça e no corpo e lobos da nadadeira caudal relativamente similares no comprimento. A espécie nova difere ainda da simpátrica H. chrysostiktos por ter sete raios ramificados na nadadeira dorsal ao invés de 10-11 e representa a décima primeira espécie de siluriforme endêmica da bacia do rio Paraguaçu.
Introduction
Hypostomus Lacépède is the second most species-rich genus of the Siluriformes (Ferraris, 2007), comprising about 130 valid species (Garavello et al., 2012; Zawadzki et al., 2012). Ten species of Hypostomus are reported from Brazilian northeastern coastal drainages and eight are known to occur in the rio São Francisco basin, the largest drainage in the region. Four species of the former group were described in the State of Bahia: Hypostomus brevicauda (Günther, 1864) and H. wuchereri (Günther, 1864), both with the vague type locality stated as "Bahia", Hypostomus unae (Steindachner, 1878) from rio Una, and the recently described H. chrysostiktos Birindelli, Lima & Zanata, 2007, endemic to the rio Paraguaçu basin. The first three of those species were described from material sent to the Natural History Museum, London, by Otto Wucherer, who most likely collected the specimens in coastal river basins south of Salvador (the Capital of the State of Bahia), as suggested by the type localities of reptiles also collected by him in the cities of Canavieiras, Nazaré, and Ilhéus (Günther, 1861, 1863; Wucherer, 1861a, 1861b, 1863a, 1863b). Particularly for Hypostomus unae, its type locality could have been one of the three distinct small coastal drainages, all named rio Una: one that drains to the coastal city of Valença, within the Recôncavo Sul basin, another that drains to the city of Una, in the south of Bahia State, and the third possibility represented by a tributary of rio Paraguaçu, on a more central portion of Bahia. The imprecise type locality, allied to the somewhat vague description, and absence of a precise diagnosis of these three species, has rendered it difficult to precisely identify Hypostomus specimens of the coastal Brazilian drainages, particularly those from Bahia State. The six remaining northeastern species of Hypostomus were described from drainages situated relatively far north of Bahia State. Among those species, H. papariae (Fowler, 1941) was described from the lago Papari, and H. pusarum (Starks, 1913) was described from the Ceará-Mirim, both in Rio Grande do Norte State. This last species has been erroneously attributed to Ceará State in the literature (e.g., Carvalho & Bockmann, 2007). The four other northeastern species from Ceará State are H. carvalhoi (Miranda Ribeiro, 1937) from rio Granjeiro, H. eptingi (Fowler, 1941) from Fortaleza, H. jaguribensis (Fowler, 1915) from rio Jaguaribe, and H. nudiventris (Fowler, 1941) from rio Choró.
Eight species of Hypostomus are known from the rio São Francisco basin and could possibly occur in rivers draining into northeastern Brazilian coastal drainages. These are H. alatus Castelnau, 1855, H. francisci (Lütken, 1874), H. garmani (Regan, 1904), H. johnii (Steindachner, 1877), H. lima (Lütken, 1874), H. macrops (Eigenmann & Eigenmann, 1888), H. subcarinatus Castelnau, 1855, and H. vaillanti (Steindachner, 1877). Most of those species were originally described from the rio das Velhas, a tributary to the upper rio São Francisco, but they were not recorded in coastal drainages of the Bahia State. In fact, a detailed comparative study is necessary to establish exactly which of these species occurs in the portion of the rio São Francisco draining Bahia. In addition to the aforementioned species, only H. scabriceps (Eigenmann & Eigenmann, 1888) is known to occur in nearby coastal drainages (rio São Mateus, Espírito Santo State).
Expeditions carried out in the last six years in all coastal drainages of Bahia, Brazil, gathered a number of specimens of Hypostomus. Detailed examination of that material has allowed the resolution of some of the alpha-taxonomy problems of the genus in that part of the country. A new species of Hypostomus, apparently endemic to the rio Paraguaçu basin, is described herein.
Material and Methods
Methodology and terminology follow Boeseman (1968), Armbruster & Page (1996), Bockmann & Ribeiro (2003), and Hollanda Carvalho & Weber (2004), with the inclusion of the following measurements: anal-fin spine length, measured from insertion to tip of the first ray; anal-fin base length; body depth at dorsal-fin origin; mouth width, taken at the level of maxillary-barbel insertions; premaxillary-ramus length, measured at the largest portions of the dentary complex sensu Adriaens et al. (2009, fig. 1b, 1c, 3e); and maxillary-barbel length, taken from the free outer portion of barbel. Plate counts and nomenclature follow Schaefer (1997), with modifications by Oyakawa et al. (2005). The following additions to plate counts were also used: paired dorsal plates between the last dorsal-fin ray base and adipose-fin (not including the pre-adipose plate); and ventral plates between the last anal-fin ray base and lower caudal-fin unbranched ray, not including the plate at the base of the rays. Standard length (SL) is expressed in mm and all other measurements are expressed as percents of standard length, with the exception of subunits of the head, which are expressed as percents of head length (HL). Measurements and counts were taken on the left side of the specimens whenever possible. Meristic data are given in the description. Asterisks indicate counts of the holotype, and the frequency of each count is given in parentheses. Counts of branchiostegal rays, vertebrae, and the relative position of dorsal and anal fins in relation to the vertebral column were examined only in cleared and stained specimens (c&s), which were prepared according to Taylor & van Dyke (1985). Osteological terminology follows Schaefer (1987). In the list of specimens examined, the total number of specimens and the size range is followed by the number and size range of the measured specimens (in parentheses), if different. All of the examined specimens are alcohol preserved, except when indicated as c&s. Institutional abbreviations are: Natural History Museum, London (BMNH), Museu de Ciências e Tecnologia, Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre (MCP), Museum of Comparative Zoology, Cambridge (MCZ), Museum National d' Histoire Naturelle, Paris (MNHN), Museu de Zoologia da Universidade de São Paulo, São Paulo (MZUSP), Núcleo de Pesquisa em Limnologia, Ictiologia e Aquicultura, Universidade Estadual de Maringá, Maringá (NUP), Universidade Federal da Bahia, Salvador (UFBA). Some specimens designated as non-types had the fins and internal organs removed by fisherman. Specimens utilized for comparative analysis are listed under 'Comparative material examined'. Comparisons with Hypostomus brevicauda and H. wuchereri were based exclusively on the type material examined.
Hypostomus jaguar, new species
Figs. 1-2
Hypostomus jaguar, holotype, MZUSP 110603, 164.8 mm SL, from the rio Paraguaçu, Bahia State, Brazil.
Holotype. MZUSP 110603, 164.8 mm SL, Brazil, Bahia, Iaçu, rio Paraguaçu at Fazenda Os Touros, 18 km from Iaçu, 12°41'10.6"S 40°07'47"W, 143 m above sea level, 8 Jun 2005, A. M. Zanata, J. L. O. Birindelli, O. T. Oyakawa, M. P. Geraldes, P. Camelier & P. Moura.
Paratypes. All from Bahia, rio Paraguaçu basin, same data as holotype, except when noticed. MZUSP 90870, 13, 68.8-175.6 mm SL. UFBA 3177, 1, 139.6 mm SL, 9 Jun 2005. UFBA 6232, 2, 123.5-138.0 mm SL, 15 May 2010, A. Zanata, R. Burger & C. P. Oliveira. MZUSP 91653, 1, 129.0 mm SL; UFBA 6457, 2, 121.7-170.2 mm SL, Itaetê, rio Una, in front of Lapa do Bode, 12°56'8.1"S 41°03'53.9"W, 308 m a.s.l., 4 Dec 2005, A. M. Zanata, P. Camelier & M. P. Geraldes. UFBA 6501, 8, 85.7-175.0 mm SL (1 c&s, 85.7 mm SL); NUP 4448, 2, 126.8-152.9 mm SL, Itaberaba, Fazenda Santo Antônio, 10 km from Vila São Vicente, out of roadway BA 242, 134 m a.s.l., 7 Jun 2005, A. M. Zanata, P. Camelier, J. O. Birindelli, O. T. Oyakawa, M. P. Geraldes & P. Moura.
Non-types in fish collections. UFBA 6502, 8 (143.0-195.0) 1 c&s ,159.1 mm SL, Itaberaba, Vila de São Vicente, bought at the market in Vila São Vicente, rio Paraguaçu basin, 3 Jun 2005.
Diagnosis.Hypostomus jaguar can be easily distinguished from the syntopic H. chrysostiktos by having seven unbranched dorsal-fin rays (vs. 10-11), large dark dots on a clear background (vs. small yellow dots on a dark background), and 25 or 26 median plates (vs. 27-29). It is distinguished from H. brevicauda and H. wuchereri by having head and trunk covered by conspicuous dark dots similar in size, ranging from half to approximately one eye diameter (vs. dots smaller than the pupil), interradial membranes of anteriormost dorsal-fin rays with one series of dots (vs. membranes with two or three series of dots), ventral surface of body comparatively weakly covered by plates, with coverage when present restricted to anterior portion of abdomen (vs. ventral surface almost or completely covered by plates, including the central area between pelvic fins). The new species differs from H. unae by having conspicuous dots on head, trunk, and fins similar in size (vs. dots on lateral of trunk inconspicuous and, when visible, much larger than those on the head and fins), pectoral spine not distinctly strong with a few elongated odontodes on its distal third (vs. pectoral spine distinctly strong with many elongated and curved odontodes on its distal half); longer dorsal fin, with tip of last ray, usually reaching preadipose plate or adipose spine when adpressed (adpressed fin 36.6-44.3% vs. 29.6-38.7% and tip of last rays not reaching adipose plate or spine), and more teeth on premaxilla (47-86, mode 70) and dentary (50-81, mode 75) (vs. 29-70 [mode 51] on premaxilla and 34-89 [mode 54] on dentary). Additionally, H. jaguar can be distinguished from its congeners from coastal northeastern rivers (H. carvalhoi, H. eptingi, H. jaguribensis, H. nudiventris, H. papariae, and H. pusarum) by a series of features, including the dark conspicuous dotted color pattern of the body and fins, ventral surface of the belly comparatively weakly covered by plates, and lobes of the caudal fin relatively similarly elongated. See 'Remarks' for additional evidence for the recognition of the new species.
Description. Standard length of examined specimens, 68.8 to 175.6 mm SL. Measurements of holotype and paratypes in Table 1. Dorsal, lateral, and ventral views in Fig. 1. Dorsal profile of body convex from snout tip to dorsal-fin origin, straight and inclined from that point to middle of caudal peduncle, and straight or somewhat concave from end of adipose-fin spine to origin of dorsalmost caudal-fin rays. Ventral profile of body straight from snout to branchial aperture, straight to somewhat convex along abdomen, and straight and inclined from urogenital opening to caudal-fin base. Body relatively depressed; greatest body width at cleithral region, progressively tapering posteriorly from cleithrum; width at cleithral region greater than head depth. Greatest body depth at dorsal-fin origin, gradually tapering to caudal peduncle. Caudal peduncle robust, slightly flattened ventrally, in cross-section roughly trapezoidal on its anterior portion and more elliptical posteriorly.
Head somewhat depressed, wide, and rounded anteriorly. Head without ridges. Posterior border of supraoccipital with small triangular process. Two plates fused to each other bordering posterior margin of supraoccipital. Interorbital region straight to somewhat convex, with lateral margins slightly elevated. Eye dorsolaterally positioned, relatively small. Dorsal iris operculum present. Mouth wide; roof and area anterior to dentary teeth covered by relatively small papillae. Lips rounded, of moderate size, occupying approximately two-thirds of ventral surface of head, its posterior border not reaching transverse line between gill openings, and without distinctly fringed edges. Inner surface of lips covered by papillae, except for smooth region just behind toothed portions. Anteriormost papillae of upper lip roundish and small, followed by patch of larger and closely positioned papillae; anteriormost papillae of lower lip roundish and relatively large, followed by patch of laterally elongated papillae; distal margins of lips with band of very small papillae, more evident on upper lip. Papillae on lower lip slightly more spaced than on upper. Teeth long, bicuspid, and curved inward distally; mesial cusp longer, approximately 2.5 to 3.0 times length of lateral cusp and curved inward, similar to Hypostomus auroguttatus (Muller & Weber, 1992: Fig. 2e). Most teeth overlap each other laterally. Premaxilla with 47-86 teeth (mode 70, holotype 67), dentary with 50 to 81 (mode 75, holotype 75). Contralateral premaxillary teeth forming relatively straight line or slightly concave arch; contralateral dentaries forming concave arch facing mouth cavity in angle of approximately 120°. Maxillary barbel short, usually similar in length to orbital diameter.
Body covered with five lateral rows of moderately spinulose dermal plates. Cheeks, snout, and rostrum completely plated, except for naked area on snout tip that continues ventrally to margin of upper lip and small areas around nares. Ornamentation of compound pterotic usually similar to remaining surface of head, but with odontodes slightly larger and more sparsely distributed. Opercle weakly armed, varying from covered with relatively small odontodes similar in size with those in nearby areas to with slightly more elongated odontodes on posterior portion of opercle. Posterior portion of preopercle surface with somewhat stronger odontodes. Cheek plates slightly evertible. Dorsal-fin base naked. Median series of plates bearing lateral-line canal. Dorsal series of plates starting posteriorly, at vertical through dorsal-fin origin; flattened between end of dorsal-fin base and adipose-fin spine. Ventral series of plates usually starting at origin or slightly posterior to origin of pelvic-fin base. Twenty one (26)* or 22(1) dorsal plates, 24(25)* or 25*(2) mid-dorsal plates, 25(7)* or 26(20) median plates, 25(3)* or 26(24) mid-ventral plates, 21(19)* or 22(8) ventral plates, 13(13) or 14(14)* plates between anal and caudal fins. Three predorsal plates (two specimens with small central plate between first two plates), 7(12)* or 8(15) plates below dorsal-fin base, 5(17) or 6(10)* paired plates between dorsal and adipose fins. One (20)*, 2(6) or 3(1) preadipose plates." aligned odontodes on plates covering most of lateral portion of trunk, forming parallel rows usually more conspicuous on posterior half of body; odontodes somewhat stronger on borders of plates of mid-ventral and ventral series. Plates without ridges, except for anterior four plates of mid-ventral series and on plates of dorsal series along dorsal-fin base to adipose fin (specimen of 68.8 mm SL without ridge on dorsal series of plates). Odontodes on head usually smaller than those on trunk and not forming rows.
Ventral surface of head and abdomen with variation on plated areas; head usually with rounded patches of plates on each side, close to branchial aperture, separated by naked central area behind lower lip; some specimens with those lateral patches reduced to few small platelets; one specimen with plates completely covering ventral surface of head, except area close to lower lip border. Anterior portion of abdomen varying from almost completely naked, with few small plates on central portion between pectoral fins, to having relatively large areas covered by plates on coracoids, central, and lateral portions close to pectoral-fin base and extending shortly posterior to it; when covered, shape of central plated area on anterior portion of abdomen rounded or roughly triangular, pointed posteriorly. Variation on anterior portion of abdomen covering apparently ontogenetic, with an almost complete lack of plates in this region in specimens up to about 110.0 mm SL, but some specimens up to 170.0 mm SL with similar condition. Posterior portion of abdomen naked; rarely with few sparse platelets on central area between pelvic-fin bases. First anal-fin pterygiophore not covered by skin in specimens about 100.0 mm SL or larger (smaller specimens with few odontodes or plate completely covered with skin).
Dorsal fin II,7, its origin situated on vertical anterior to pelvic-fin origin and approximately on midlenght of pectoral-fin spine. Tips of first and last pterygiophores of dorsal fin lying above neural spines of vertebrae 7 and 16, respectively. Dorsal fin higher than length of its base; tips of adpressed last rays reaching anterior border of preadipose plate or, more rarely, the anterior border of adipose-fin spine. Posterior margin of dorsal fin slightly convex. Adipose-fin spine narrow, usually straight or slightly curved ventrally. Pectoral fin I,6, its posterior margin slightly concave; pectoral-fin spine not distinctly strong. Distal portion of unbranched pectoral-fin ray with odontodes somewhat longer and slightly curved, more conspicuous on larger individuals. Degree of overlap of adpressed pectoral-fin tip on pelvic fin ontogenetically variable, with tip of fin reaching mid-length of unbranched pelvic-fin ray in smaller and medium-sized specimens and reaching posterior third of pelvic fin in largest individuals. Pectoral fin inserted slightly dorsally to level of pelvic-fin insertion and pectoral-fin spine resting on top of pelvic fin when adpressed. Pelvic fin i,5, its posterior margin somewhat rounded to straight. Tip of pelvic fin usually reaching to anterior one fourth to half of anal-fin unbranched ray. Anal fin i,3(1) or i,4(27)*, its border straight to slightly rounded and reaching fifth or sixth plate after its origin. Tips of first and last pterygiophores of anal fin lying below haemal spines of vertebrae 14 and 17, respectively. Caudal fin i,14,i(25)* or i,15,i(2), its posterior margin varying from somewhat concave to straight; lower unbranched ray somewhat longer than upper. All fin rays covered by odontodes, somewhat stronger on unbranched rays. Twenty nine vertebrae, three branchiostegal rays (1).
Color in alcohol. Overall ground color of dorsal and lateral surface of body yellowish to light brown. Dorsal and lateral surface of body covered with large black, conspicuous, and equally distributed dots. Dots on trunk usually varying from half to approximately an eye diameter, decreasing gradually in size towards head. Diameter of dots on head usually about half eye diameter, slightly smaller on snout. Fins covered with similar dots, slightly smaller than the ones on trunk and slightly larger than most dots on head. Interradial membranes of anteriormost dorsal-fin rays with one series of dots and posteriormost interradial membranes with two series; few specimens with two or three dots elongated and merged with each other, forming short longitudinal or inclined bands; membrane between longest rays with seven to nine dots, and membrane between last two rays with vertical line of four or five dots in specimens around 80.0 mm SL or larger; number of dots on membranes sometimes lower in smaller specimens. Pectoral and pelvic fins similarly dotted, but dots not fused and not forming bands; spines usually with seven or more dots in specimens around 80.0 mm SL or larger, and four or five on smaller. Adipose spine with three dots; adipose membrane dotted or plain. Caudal fin with dots on rays and membranes, usually not forming vertical stripes, but in some specimens (usually small ones) dots somewhat merged to each other forming weakly defined vertical stripes; appearance of vertical stripes observed also in some specimens when caudal fin somewhat folded. Around 10 dots on unbranched caudal-fin rays and eight or more on the longest branched rays. Anal fin dotted. All fins with posterior margins darkened. Overall ground color of ventral body surface yellowish or light brown; black round dots usually absent, when present concentrated on head, on plated portion, or sparsely distributed on belly. Dots on belly, when present, less conspicuous than on lateral portion of body. Ventral surface of caudal peduncle yellowish, usually without conspicuous dots, with darkened median longitudinal stripe or few sparse darkened roundish areas. Largest specimen examined darker overall, with somewhat grayish background and dots slightly less visible, although with pattern described above.
Color in life. Description based on field observations of several specimens and on pictures of living specimens (Fig. 2). Ground color light brown or yellowish with somewhat golden hue. Black, round, and conspicuous dots on body and fin rays and interradial membranes similar to coloration in alcohol-preserved specimens. Ventral surface of head and abdomen yellowish to light brown, with or without dots.
Etymology. From îagûara in the indigenous language Tupi-Guarani, which means panther, in allusion to the black dotted color pattern on a clear background. A noun in apposition.
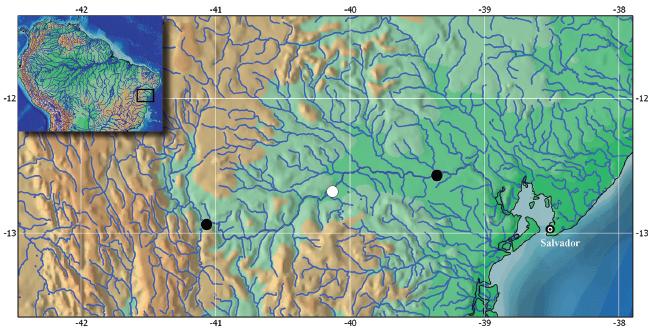
Distribution. The new species is known from the rio Paraguaçu basin (Fig. 3), an independent coastal drainage in northeastern Brazil situated on the eastern slopes of the Chapada Diamantina, Bahia State.
Habitat notes. Specimens of Hypostomus jaguar were captured in the rio Paraguaçu and its tributaries, in stretches of up to 60 m wide and 1.5 m deep, with rocky bottom, dark colored water, mild to fast water current, and at altitudes ranging from 143 to 350 meters above sea level. The riparian vegetation of the rio Paraguaçu basin is highly variable, including domains such as "Campo Rupestre", "Caatinga", "Cerrado" (Brazilian savannas) and semi-deciduous seasonal to evergreen forests. The habitats in most areas sampled were anthropized mainly due to cattle ranches. The species occurs sympatrically with Hypostomus chrysostiktos.
Common name. Acari, acari-preto, acari-pintado.
Remarks. In addition to the overall well-defined large dotted color pattern of Hypostomus jaguar, contrasting with the small to moderate sized and usually faded dots on body and fins coloration of the northeastern non-Bahian congeners (H. carvalhoi, H. eptingi, H. jaguribensis, H. nudiventris, H. papariae, H. pusarum), it also differs by having the ventral surface of body naked or weakly plated, always without a broad plated area on posterior portion of abdomen (vs. broad plated area on anterior and posterior portion of abdomen of H. eptingi, and anterior portion of abdomen completely covered by small plates forming a broad rectangular area continuing posteriorly through a narrow median longitudinal band, followed by a somewhat broad plated area situated in the region posterior to pelvic-fin base in H. carvalhoi, H. jaguribensis, H. papariae, and H. pusarum). Hypostomus jaguar differs further from the last four species by the absence of ridges on trunk (vs. presence). The absence of dark bands on the caudal fin further distinguishes H. jaguar from H. eptingi and H. nudiventris, with five transverse dark bands on the former species and seven on the latter. The new species also differs from H. eptingi by having the lobes of the caudal fin relatively similar in length (vs. lower lobe distinctly longer than upper) and from H. nudiventris by its dorsal-fin interradial membrane coloration (absence of brown median vertical short lines or streaks parallel to rays vs. its presence).
Eight species of Hypostomus are known from the rio São Francisco basin (H. alatus, H. francisci, H. garmani, H. johnii, H. lima, H. macrops, H. subcarinatus, and H. vaillanti). As previously noted above, none of these species have so far been recorded in the coastal rivers of the Bahia State and none of them were identified among specimens collected in the rio Paraguaçu basin examined in this study. All of the Hypostomus species described from the rio São Francisco basin have the ventral surface of the head and trunk almost or completely covered by plates, in contrast to a naked or weakly covered ventral surface of the belly in the species described herein. In addition, H. jaguar is readily distinguished from H. alatus and H. francisci by having dark dots against a yellowish or clear brown background (vs. light spots against a dark background), from H. johnii and H. vaillanti by having caudal-fin lobes relatively similar in length (vs. lower lobe distinctly longer than upper), from H. subcarinatus by having a robust body and absence of predorsal ridges (vs. elongate body and at least three predorsal ridges), from H. macrops by having the pectoral spine weakly armed with odontodes (vs. pectoral spine with large and curved odontodes on most of its extension), and from H. lima by having dots similar in size along the head and trunk (vs. spots on the trunk twice as large as those on the head). From H. scabriceps, the only congener known from coastal drainages nearby, to the south of Bahia State, the new species can be further distinguished by having one or two dots covering each body plate (vs. comparatively small dots, up to four dots per plate) and dorsal fin reaching to or almost reaching to the adipose fin (vs. dorsal fin distant by at least two plates from the adipose fin).
In addition, compared with other congeners, H. jaguar can be distinguished from species of the H. cochliodon group, with the exception of H. hemicochliodon, by having bifid non-spoon-shaped teeth and the mesial and lateral cusps not being fused (vs. spoon-shaped teeth with mesial and lateral cusps usually fused). Hypostomus jaguar can be distinguished from H. alatus, H. albopunctatus, H. faveolus, H. francisci, H. lexi, H. luteus, H. luteofrenatus, H. margaritifer, H. microstomus, H. multidens, H. regani, H. roseopunctatus, H. scaphiceps, H. strigaticeps, H. tietensis, and H. variipictus by having conspicuous black dots on the body and fins (vs. pale or yellowish spots or blotches). The new species is further distinguished from the remaining Hypostomus species, except H. agna, H. brevicauda, H. brevis, H. garmani, H. heraldoi, H. hermanni, H. johnii, H. lima, H. luetkeni, H. nigropunctatus, and H. wuchereri, by the absence of ridges on the compound pterotic, predorsal plates and lateral series of plates (vs. having moderate to developed ridges on the compound pterotic, predorsal plates, and lateral series of plates). The new species differs further from H. agna, H. brevicauda, H. heraldoi, H. johnii, H. luetkeni, H. nigropunctatus, H. vermicularis, and H. wuchereri by the head and trunk being covered by large and conspicuous dark dots, the diameter of dots ranging from half to approximately one eye diameter (vs. small dots, similar in diameter to the pupil diameter or smaller); and from H. garmani, H. hermanni, H. lima, and H. mutucae by having the ventral surface of the body comparatively weakly covered by plates, that are restricted, when present, to the portion between the pectoral-fin bases (vs. ventral surface almost or completely covered by plates, including the median area between the pelvic fins).
Hypostomus jaguar is one of the two species of the genus that is endemic to the rio Paraguaçu basin. Its sympatric congener, H. chrysostiktos, has a higher number of branched dorsal-fin rays (10-11), which is a remarkable and unique characteristic within the Hypostomini, together with an inverted pattern of coloration. Thus, H. jaguar has external morphological features that are apparently more similar to congeners from other drainages (e.g., H. unae, known to the rio Una, rio Jequiriçá and rio das Almas basins) than with the sympatric H. chrysostiktos. Although a study of relationships was not performed, we suggest that the closest relatives of H. jaguar are possibly congeners from outside the rio Paraguaçu basin. The endemic nature of the rio Paraguaçu ichthyofauna has been previously highlighted (e.g., Lima & Gerhard, 2001; Santos & Zanata, 2006; Birindelli et al., 2007; Santos & Caramaschi, 2007) and a total of 16 described species are now known to be endemic to the basin. Hypostomus jaguar represents the tenth siluriform species that is apparently endemic to the Paraguaçu basin.
Weber (2003) mentioned the possibility that H. wuchereri, a species with the vague type locality "Bahia", occurred in the rio Paraguaçu, as previously cited by Bitencourt et al. (2011). However, the examination of some of the type material of H. wuchereri revealed that it is a distinct species from H. chrysostiktos and H. jaguar, the two species that are known to occur in the rio Paraguaçu basin. Extensive collections in the rio Paraguaçu in the last 12 years failed to capture specimens of H. wuchereri and, consequently, the occurrence of the species in this river basin seems unlikely. A taxonomic study aiming to redescribe H. wuchereri and define its distribution is currently under way by the authors.
Comparative material examined. Brazil:Hypostomus brevicauda, BMNH 1864.1.19.16 -17, 2 syntypes, 189.0-196.1 mm SL, Bahia. Hypostomus chrysostiktos, UFBA 2786, 5 paratypes, 112.3-259.7 mm SL, Bahia, Iaçu, rio Paraguaçu. Hypostomus garmani, BMNH 1904.1.28.3, holotype, 209.9 mm SL, Minas Gerais, rio das Velhas. Hypostomus heraldoi, MZUSP 98771, holotype, 217.9 mm SL, Goiás, rio Pirapitinga. Hypostomus johnii, MCZ 7831, 1 syntype, 94.0 mm SL, Piauí, Teresina, rio Parnaíba basin, rio Poti. MCZ 7864, 2 syntypes, 93.1-95.5 mm SL, Piauí, Teresina, rio Parnaíba basin, rio Poti. Hypostomus kuarup, NUP 9145, 5, 126.8-148.8 mm SL; NUP 9146, 10, 98.0-152.6 mm SL; NUP 9200, 17, 32.0-117.6 mm SL, Mato Grosso, Campinápolis, rio Culuene. Hypostomus lima, BMNH 1876.1.10, 2 syntypes, 72.9-86.1 mm SL, Minas Gerais, Lagoa Santa. Hypostomus multidens, NUP 5340, paratype, 157.0 mm SL, São Paulo, rio Paraná basin, rio Paranapanema. NUP 5340, 1, 157.0 mm SL, São Paulo, Piraju, upper rio Paraná basin, rio Paranapanema, Chavantes reservoir. NUP 6776, 1, 167.0 mm SL, Paraná, Doutor Oliveira Castro, rio Paraná, upper rio Paraná basin. Hypostomus mutucae, MCP 28669, holotype, 67.7 mm SL, Mato Grosso, Chapada dos Guimarães, rio Paraguai basin, rio Mutuca. MZUSP 27694, 2, 75.0-79.4 mm SL, Mato Grosso, Chapada dos Guimarães, rio Paraguai basin, rio Mutuca. NUP 6641, 13, 52.4-109.2 mm SL, Mato Grosso, Chapada dos Guimarães, rio Paraguai basin, rio Claro. NUP 6642, 4, 62.1-98.1 mm SL, Mato Grosso, Chapada dos Guimarães, rio Paraguai basin, rio Claro. Hypostomus nigropunctatus, NUP 5081, paratype, 84.3 mm SL; NUP 5082, 1 paratype, 171.4 mm SL; NUP 5083, paratype, 208.4 mm SL, Paraná, rio Iguaçu basin. Hypostomus pusarum, UFBA 3824, 3, 84.2-107.9 mm SL, Rio Grande do Norte, Jardim do Seridó, rio Seridó. Hypostomus scabriceps, BMNH 1904.1.28.3, 1 syntype, 88.0 mm SL, Espírito Santo, São Matheos. Hypostomus strigaticeps BMNH 1907.7.6.1012, 3 syntypes, 75.7-160.0 mm SL, São Paulo, rio Tietê basin, rio Piracicaba. NUP 4017, 2, 72.8-100.0 mm SL, São Paulo, Ipuã, rio Tietê basin, rio Ipanema. NUP 4538, 11, 82.0-140 mm SL, São Paulo, Piracicaba, rio Tietê basin, rio Corumbataí. Hypostomus subcarinatus, MNHN A.9575, holotype, 241.8 mm SL, Minas Gerais. Hypostomus wuchereri, BMNH1863.3.27.15, 1 syntype, 203.8 mm SL, Bahia. BMNH 1852.13.12.8, 1 syntype, 127.3 mm SL, Bahia. Hypostomus sp., UFBA 3027, 5, 115.0 -123.0 mm SL, Bahia, Rio Real, rio Itapicuru. Hypostomus sp., UFBA 4254, 3, 83.5-79.4 mm SL, Bahia, Floresta Azul, rio Salgado, rio Cachoeira basin. Hypostomus sp., UFBA 4835, 1, 155.0 mm SL, Bahia, Mascote, rio Pardo. Hypostomus sp., UFBA 5862, 2, 22.3-40.0 mm SL, Bahia, Muniz Ferreira, rio da Onha, rio Jaguaripe basin. Hypostomus sp., UFBA 6335, 4, 60.4-174.0 mm SL, Bahia, Ubaitaba, rio Oricó, tributary of rio de Contas. Hypostomus sp., UFBA 6457, 2, 123.0-170.0 mm SL, Bahia, Itaetê, rio Una, tributary of rio Paraguaçu. Hypostomus sp., NUP 9813, 13, 51.6-76.8 mm SL, Bahia, Jequié, rio das Pedras, tributary of rio das Contas.
The manuscript benefited from comments and suggestions of M. R. Britto (MNRJ) and O. T. Oyakawa (MZUSP). We thank A. Góes, C. P. Oliveira, J. L. Birindelli, M. P. Geraldes, O. T. Oyakawa, P. Moura, P. Camelier, and R. Burger for help in the fieldwork. A photograph of a preserved specimen was provided by Eduardo Baena. Priscila Camelier also helped with the map. For the loan of specimens and other assistance we thank C. Lucena (MCP), O. Oyakawa (MZUSP), and R. S. Rosa (UFPB). Recent material was acquired in field expeditions funded by All Catfish Species Inventory (NSF DEB-0315963), during an expedition to Paraguaçu and Itapicuru rivers (EPI), in July 2005, and CNPq (# 133917/2010-0) on other occasions. CHZ had grants from CNPq, Proc. 306066/2009-2. BSB had a master fellowship from CNPq. Permission for collecting specimens was granted by IBAMA and ICMBio.
Literature Cited
- Adriaens, D., T. Geerinckx, J. Vlassenbroeck, L.van Hoorebeke & A. Herrel. 2009. Extensive jaw mobility in suckermouth armored catfishes (Loricariidae): a morphological and kinematic analysis of substrate scraping mode of feeding. Physiological and Biochemical Zoology, 82: 51-62.
- Alves, C. B. M. & P. S. Pompeu. 2001. Peixes do Rio das Velhas: passado e presente. SEGRAC. Belo Horizonte.
- Armbruster, J. W. & L. Page. 1996. Redescription of Aphanotorulus (Teleostei: Loricariidae) with description of one new species, A. ammophilus, from the Río Orinoco Basin. Copeia, 1996: 379-389.
- Birindelli, J. L., A. M. Zanata & F. C. T. Lima. 2007. Hypostomus chrysostiktos, a new species of armored catfish (Siluriformes: Loricariidae) from rio Paraguaçu, Bahia State, Brazil. Neotropical Ichthyology, 5: 271-278.
- Bitencourt, J. A., P. R. A. M. Affonso, L. Giuliano-Caetano & A. L. Dias. 2011. Identification of distinct evolutionary units in allopatric populations of Hypostomus cf. wuchereri Günther, 1864 (Siluriformes: Loricariidae): karyotypic evidence. Neotropical Ichthyology, 9: 317-324.
- Bockmann, F. A. & A. C. Ribeiro. 2003. Description of a new suckermouth armored catfish of the genus Pareiorhina (Siluriformes: Loricariidae) from southeastern Brazil. Ichthyological Exploration of Freshwaters, 14: 231-242.
- Boeseman, M. 1968. The genus Hypostomus Lacépède, 1803, and its Surinam representatives (Siluriformes, Loricariidae). Zoologische Verhandelingen, 99: 1-9.
- Carvalho, M. & F. A. Bockmann. 2007. Família Loricariidae: Hypostominae. In: Buckup, P. A., Menezes, N. A. & Ghazzi, M. S. (Eds.). Catálogo das espécies de peixes de água doce do Brasil. Série livros 23, Museu Nacional, Rio de Janeiro.
- Castelnau, F. L. 1855. Poissons. In: Castelnau, F. L. Animaux nouveaux or rares recueillis pendant l'expédition dans les parties centrales de l'Amérique du Sud, de Rio de Janeiro a Lima, et de Lima au Para. Bertrand, Paris, 2.
- Eigenmann, C. H. & R. S. Eigenmann. 1888. Preliminary notes on South American Nematognathi. Proceedings of the California Academy of Sciences (Series 2), 2: 28-56.
- Ferraris Jr., C. J. 2007. Checklist of catfishes, recent and fossil (Osteichthyes: Siluriformes), and catalogue of siluriform primary types. Zootaxa, 1418: 418-628.
- Fowler, H. W. 1915. Notes on nematognathous fishes. Proceedings of the Academy of Natural Sciences of Philadelphia, 67: 203-243.
- Fowler, H.W. 1941. A collection of fresh-water fishes obtained in eastern Brazil by Dr.Rodolpho von Ihering. Proceedings of the Academy of Natural Sciences of Philadelphia, 93: 123-199.
- Garavello, J. C., H. A. Britski & C. H. Zawadzki. 2012. The cascudos of the genus Hypostomus Lacépède (Ostariophysi: Loricariidae) from the rio Iguaçu basin. Neotropical Ichthyology, 10: 263-283.
- Günther, A. 1861. Account of the reptiles sent by Dr. Wucherer from Bahia. Proceedings of the Zoological Society of London, 1861: 12-18.
- Günther, A. 1863. Addition to Dr. Wucherer's article on the ophidians of Bahia. Proceedings of the Zoological Society of London, 1863: 56-58.
- Günther, A. 1864. Catalogue of the fishes in the British Museum. Vol. 5. Catalogue of the Physostomi, containing the families Siluridae, Characinidae, Aplochitonidae, Sternoptychidae, Scopelidae, Stomiatidae in the collection of the British Museum. London, Taylor and Francis.
- Hollanda Carvalho, P. & C. Weber. 2004. Five new species of the Hypostomus cochliodon group (Siluriformes:Loricariidae) from the middle and lower Amazon System. Revue suisse de Zoologie, 4: 953-978.
- Lima, F. C. T. & P. Gerhard. 2001. A new Hyphessobrycon (Characiformes: Characidae) from Chapada Diamantina, Bahia, Brazil, with notes on its natural history. Ichthyological Exploration of Freshwaters, 12: 105-114.
- Miranda Ribeiro, A. 1937. Sobre uma collecção de vertebrados do nordeste brasileiro. Primeira parte: Peixes e batrachios. O Campo, 1: 54-56.
- Muller, S. & C. Weber. 1992. Les dents des sous-familles Hypostominae et Ancistrinae (Pisces, Siluriformes, Loricariidae) et leur valeur taxonomique. Revue suisse de Zoologie, 99: 747-754.
- Oyakawa, O. T., A. Akama & A. M. Zanata. 2005. Review of the genus Hypostomus Lacépède, 1803 from rio Ribeira do Iguape basin, with description of a new species (Pisces, Siluriformes, Loricariidae). Zootaxa, 921: 1-27.
- Regan, C. T. 1904. A monograph of the fishes of the family Loricariidae. Transactions of the Zoological Society of London, 17: 191-168.
- Santos, A. C. A. & A. M. Zanata. 2006. Peixes no semi-árido brasileiro, p. 97-102. In: Queiroz, L. P., A. Rapini & A. M. Giulietti (Eds). Rumo ao Amplo Conhecimento da Biodiversidade do Semi-árido Brasileiro. Brasília, Ministério da Ciência e Tecnologia.
- Santos, A. C. A. & E. P. Caramaschi. 2007. Composition and seasonal variation of the ichthyofauna from upper rio Paraguaçu (Chapada Diamantina, Bahia, Brazil). Brazilian Archives of Biology and Technology, 50: 663-672.
- Schaefer, S. A. 1987. Osteology of Hypostomus plecostomus (Linnaeus) with phylogenetic analysis of the loricariid subfamilies (Pisces: Siluroidei). Contributions in Sciences, Natural History Museum of Los Angeles County, 394: 1-31.
- Schaefer, S. A. 1997. The Neotropical cascudinhos: systematics and biogeography of the Otocinclus catfishes (Siluriformes, Loricariidae). Proceedings of the Academy of Natural Sciences of Philadelphia, 148: 1-120.
- Starks, E. C. 1913. The fishes of the Stanford Expedition to Brazil. Leland Stanford Junior University Publications, University Series, 15: 1-77.
- Steindachner, F. 1878. Ichthyologische Beiträge. VI. Sitzungsberichten der Kaiserlichen Akademie der Wissenschaften in Wien, 77: 379-392.
- Taylor, W. R. & G.C. vanDyke. 1985. Revised procedures for staining and clearing small fishes and other vertebrates for bone and cartilage study. Cybium, 9: 107-119.
- Zawadzki, C. H., J. L. O. Birindelli & F. C. T. Lima. 2012. A new armored catfish species of the genus Hypostomus Lacépède, 1803 (Siluriformes: Loricariidae) from the upper rio Xingu basin, Brazil. Neotropical Ichthyology, 10: 245-253.
- Weber, C. 2003. Subfamily Hypostominae (Armored catfishes), p. 351-372. In: Reis, R. E., S. O. Kullander & C. J. Ferraris (Eds.). Check List of the Freshwater Fishes of South and Central America. Porto Alegre, Edipurcs, Porto Alegre.
- Wucherer, O. 1861a. On the ophidians of the Province of Bahia, Brazil. Proceedings of the Zoological Society of London, 1861: 113-117.
- Wucherer, O. 1861b. Description of a new species of Elapomorphus from Brazil. Proceedings of the Zoological Society of London, 1861: 325-326.
- Wucherer, O. 1863a. On the species of Craspedocephalus which occur in the Province of Bahia. Proceedings of the Zoological Society of London, 1863: 51-54.
- Wucherer, O. 1863b. On the ophidians of the Province of Bahia, Brazil. Proceedings of the Zoological Society of London, 1863: 55-56.
-
Published June 28, 2013
Publication Dates
-
Publication in this collection
June 2013
History
-
Received
03 Aug 2012 -
Accepted
20 Mar 2013