
Abstracts
Plant population and arrangement should be properly designed to maximize the seed yield of short-height genotypes of castor (Ricinus communis L.) in each cropping season. Experiments were performed in the in the fall-winter cropping season of 2008 and 2009 in Botucatu, State of São Paulo, Brazil, to evaluate the effects of row spacing (0.45, 0.60, 0.75, and 0.90 m) and plant population (25,000, 40,000, 55,000, and 70,000 plants ha-1) on agronomical traits, yield components, seed yield, and oil yield of the genotype FCA-PB. A high plant density caused a reduction in plant survival and increased the first raceme insertion height, particularly in the year with the higher rainfall. The basal stem diameter, number of racemes per plant, and seeds per raceme were reduced by increasing the plant density. However, the seed and oil yields were minimally influenced by plant population density and row spacing because the castor plant compensated for a low plant population with a high number of racemes and seeds. The maximum seed yield was achieved with an estimated 0.64 m of row spacing and 50,107 plants ha-1 (i.e., 3.2 plants m-1).
Ricinus communis; spatial plant distribution; plant density; grain yield; oil content
Para maximizar a produtividade de grãos de genótipos de mamona (Ricinus communis L.) de porte baixo, devem ser definidos a população e o arranjo de plantas mais adequados a cada época de cultivo. Foi conduzido um experimento no outono-inverno dos anos de 2008 e 2009, em Botucatu-SP, para avaliar os efeitos do espaçamento entre fileiras (0,45; 0,60; 0,75 e 0,90 m) e da população de plantas (25.000, 40.000, 55.000 e 70.000 plantas ha-1) sobre as características agronômicas, componentes da produção, produtividade de grãos e de óleo do genótipo FCA-PB. Elevada densidade de plantas provocou redução na sobrevivência das mesmas e aumentou a altura de inserção do primeiro racemo, especialmente no ano com maior disponibilidade hídrica. O diâmetro do caule, o número de racemos por planta e de sementes por racemo foram reduzidos com o aumento da densidade de plantas. As produtividades de grãos e de óleo foram pouco influenciadas pela população de plantas e espaçamento entre fileiras, pois a mamoneira compensa um baixa população de plantas, com um elevado número de racemos e grãos por planta. A máxima produtividade de grãos foi obtida com o espaçamento entre fileiras estimado de 0,64 m e 50.107 plantas ha-1 (ou seja, 3,2 plantas m-1).
Ricinus communis; distribuição especial de plantas; densidade de plantas; produtividade de grãos; teor de óleo
PRODUÇÃO VEGETAL/CROP PRODUCTION
Optimizing row spacing and plant population arrangement for a new short-height castor genotype in fall-winter
Combinação ótima de espaçamento e população de plantas para um novo genótipo de mamona de porte baixo na safrinha
Genivaldo David de Souza-SchlickI; Rogério Peres SorattoII, * * Author for correspondence. E-mail: soratto@fca.unesp.br ; Mauricio Dutra ZanottoII
IPrograma de Pós-graduação em Agronomia-Agricultura, Faculdade de Ciências Agronômicas, Universidade Estadual Paulista, Botucatu, São Paulo, Brazil
IIDepartamento de Produção e Melhoramento Vegetal, Faculdade de Ciências Agronômicas, Universidade Estadual Paulista, Rua José Barbosa de Barros, 1780, 18610-307, Botucatu, São Paulo, Brazil
ABSTRACT
Plant population and arrangement should be properly designed to maximize the seed yield of short-height genotypes of castor (Ricinus communis L.) in each cropping season. Experiments were performed in the in the fall-winter cropping season of 2008 and 2009 in Botucatu, State of São Paulo, Brazil, to evaluate the effects of row spacing (0.45, 0.60, 0.75, and 0.90 m) and plant population (25,000, 40,000, 55,000, and 70,000 plants ha-1) on agronomical traits, yield components, seed yield, and oil yield of the genotype FCA-PB. A high plant density caused a reduction in plant survival and increased the first raceme insertion height, particularly in the year with the higher rainfall. The basal stem diameter, number of racemes per plant, and seeds per raceme were reduced by increasing the plant density. However, the seed and oil yields were minimally influenced by plant population density and row spacing because the castor plant compensated for a low plant population with a high number of racemes and seeds. The maximum seed yield was achieved with an estimated 0.64 m of row spacing and 50,107 plants ha-1 (i.e., 3.2 plants m-1).
Keywords:Ricinus communis, spatial plant distribution, plant density, grain yield, oil content.
RESUMO
Para maximizar a produtividade de grãos de genótipos de mamona (Ricinus communis L.) de porte baixo, devem ser definidos a população e o arranjo de plantas mais adequados a cada época de cultivo. Foi conduzido um experimento no outono-inverno dos anos de 2008 e 2009, em Botucatu-SP, para avaliar os efeitos do espaçamento entre fileiras (0,45; 0,60; 0,75 e 0,90 m) e da população de plantas (25.000, 40.000, 55.000 e 70.000 plantas ha-1) sobre as características agronômicas, componentes da produção, produtividade de grãos e de óleo do genótipo FCA-PB. Elevada densidade de plantas provocou redução na sobrevivência das mesmas e aumentou a altura de inserção do primeiro racemo, especialmente no ano com maior disponibilidade hídrica. O diâmetro do caule, o número de racemos por planta e de sementes por racemo foram reduzidos com o aumento da densidade de plantas. As produtividades de grãos e de óleo foram pouco influenciadas pela população de plantas e espaçamento entre fileiras, pois a mamoneira compensa um baixa população de plantas, com um elevado número de racemos e grãos por planta. A máxima produtividade de grãos foi obtida com o espaçamento entre fileiras estimado de 0,64 m e 50.107 plantas ha-1 (ou seja, 3,2 plantas m-1).
Palavras-chave:Ricinus communis, distribuição especial de plantas, densidade de plantas, produtividade de grãos, teor de óleo.
Introduction
Castor (Ricinus communis L.) oil is distinct from other vegetable oils, mainly because it consists of up to 90% of a hydroxylated fatty acid called ricinoleic acid (SEVERINO et al., 2012), and it has many applications in the chemical industry (BARNES et al., 2009), including biodiesel production (BALDWIN; COSSAR, 2009).
The castor plant is tolerant to drought and adapted to many cropping conditions (BABITA et al., 2010; CARVALHO et al., 2010; ZHOU et al., 2010). In Brazil, new castor genotypes have been developed with a high yield potential, a short growing cycle, even maturation, indehiscent fruit, and a short stature and are suitable for mechanical harvesting techniques. Furthermore, castor is an attractive option for growth as a second crop (out-of-season harvest) in the fall-winter cropping season, particularly in succession to soybean [Glycine max (L.) Merr.]. In fall-winter, traditional crops, such as corn (Zea mays L.) and sunflower (Helianthus annuus L.), are not grown because rain is scarce and erratic (SAVY FILHO, 2005; SILVA et al., 2010).
The optimization of row spacing and in-row plant density is a simple procedure with a low cost but has a significant influence on yield (SEVERINO et al., 2006a and b; SEVERINO et al., 2012; SORATTO et al., 2012) and is essential to maximize seed production (COX; CHERNEY, 2011). A high plant density may result in overgrown plants (CARVALHO et al., 2010) and subsequent lodging, whereas a low plant population may favor weed infestation, late flowering, long lateral branches, and wide stems, which impair mechanical harvesting (LOPES et al., 2008; SEVERINO et al., 2006b, 2012). Light interception by plants strongly influences the crop yield when other environmental factors are favorable, and it is modified by the plant spatial distribution in a given area (SEVERINO et al., 2006a and b; 2012).
Nevertheless, the optimum plant arrangement depends on the characteristics of each genotype, such as the growth habit, plant stature and architecture (SEVERINO et al., 2006a and b), and environmental conditions (SEVERINO et al., 2006a; SORATTO et al., 2012). Kittock and Williams (1970) observed that, for short-height castor in a 1.02 m row spacing, the best plant population was 58,000 plants ha-1 under irrigation and 30,000 plants ha-1 under rainfed conditions. For the IAC 2028 cultivar, plant populations greater than 25,000 plants ha-1 did not increase the seed yield, regardless of the row spacing used, in the fall-winter cropping season (SOUZA-SCHLICK et al., 2011). Soratto et al. (2012) reported that the seed yield of castor genotypes in fall-winter was greater for more equidistant plant arrangements and that an increase in the plant population did not affect the seed yield when the same row spacing (0.90 m in this case) was used. However, there are no conclusive studies and recommendations to date for the optimum row spacing and plant population arrangement for the growth of new short-height castor genotypes as a second crop in fall-winter under a mechanized cropping system.
This study aimed to evaluate the effects of row spacing and plant population arrangement on agronomical traits, yield components, seed yield, and oil yield of the short-height castor genotype FCA-PB grown in the fall-winter season.
Material and methods
The experiment was performed in Botucatu, São Paulo State, Brazil (48o 23' W; 22o 51' S; 740 m above sea level), during the fall-winter cropping season 2008 and 2009. The soil was a sandy clay loam, kaolinitic, thermic Typic Haplorthox (distroferric Red Latosol). The chemical characteristics of the topsoil (0-0.20 m) were determined at the beginning of each experiment. In 2008, the soil characteristics were as follows: pH (CaCl2) of 5.0; 38 g dm-3 of total organic matter; 39.0 mg dm-3 of p (resin); 4.1, 31.0, 9.0, and 51.0 mmolc dm-3 of exchangeable K, Ca, Mg, and H+Al, respectively; and base saturation of 46%. The values in 2009 were as follows: pH (CaCl2) of 5.6; 40 g dm-3 of total organic matter; 26.0 mg dm-3 of p (resin); 2.6, 43.0, 22.0, and 29.0 mmolc dm-3 of exchangeable K, Ca, Mg, and H+Al, respectively; and base saturation of 70%.
The climate of the region is Cwa according to the Köppen classification, which is characterized as a tropical, with a dry winter and a hot rainy summer. The data for the rainfall and temperature registered during the experiments are presented in Figure 1.
The experimental design was a completely randomized block in a split-plot arrangement, which was replicated four times, with the row spacings assigned to the main plots and the plant populations assigned to the subplots. The treatments were a factorial distribution of four row spacings (0.45, 0.60, 0.75, and 0.90 m) and four plant populations (25,000, 40,000, 55,000, and 70,000 plants ha-1) as the subplots. The main plots were 20 m long and consisted of ten rows when the spacing was 0.45 m and six rows when the spacing was 0.60, 0.75, or 0.90 m. The subplots were 5 m long. Data were not obtained for the plants at 0.5 m at the end of each row and in the lateral rows of each subplot.
The FCA-PB genotype was developed in the breeding program of the College of Agricultural Sciences - São Paulo State University (UNESP). Its characteristics are a short, indehiscent fruits, susceptibility to gray mold [Botryotinia ricini (G.H. Godfrey) Whetzel], an oil content higher than 47%, and a short cycle (130-150 days).
A no-tillage system was adopted. Two weeks prior to sowing, the area was sprayed with 1.44 kg a.i. ha-1 of glyphosate. Furrow opening and fertilizer distribution was performed using a sowing-fertilizer machine; mineral fertilizer was applied at a dose of 150 kg ha-1 of NPK 8-28-16. Sowing was manually performed in the first half of March 2008 and 2009 using an excessive number of seeds. The seeds were placed at a 4 cm depth and covered with soil. Seedling emergence occurred 10 days after sowing, and thinning was performed at 10 days after emergence (DAE) to achieve the assigned plant population. Topdressing fertilization using 60 kg ha - 1 of N (ammonium sulfate) was performed at 30 DAE.
Weeding was performed by hand. Flowering of the first raceme occurred at 42 DAE in 2008 and 59 DAE in 2009. After the beginning of flowering, fungicides were sprayed to prevent gray mold infestation. In 2008, procymidone at 0.5 kg a.i. ha-1 was sprayed three times, and iprodione at 0.5 g i.a. ha-1 was sprayed once. The plants were sprayed with 0.5 kg ha-1 of procymidone four times in 2009.
The plants were harvested at 153 DAE in 2008 and 185 DAE in 2009. The plant survival rate was calculated for all the plants in two 4-m-long rows. The basal stem diameter (at the first internode), plant height (distance from the soil to the highest point of the plant), and first raceme insertion height (distance from the soil to the point of raceme insertion) were measured for ten plants in each subplot. All the racemes in two 4-m-long rows were hand-harvested. The fruits were counted and hand-threshed, and the seeds were counted and weighed; the seed yield was adjusted to 130 g kg-1 of moisture content.
The seeds were oven dried at 70°C for 24 hours to maximize the sample homogeneity. A subsample (12 to 15 g) from each plot was weighed, and the seed oil content was determined using a bench-top NMR spectrometer model SLK-100 (SpinLock, Cordoba, Argentina), following the method described by Colnago et al. (2007). The oil content (%) was expressed on a seed dry matter basis.
The Lilliefors test was used to verify the normality of the data. The homogeneity of the variances was evaluated using the Cochran-Bartlett test. The data were subjected to an ANOVA using a split-plot scheme in the statistical program SISVAR. The blocks and all the block interactions were considered random effects; the year, row spacing, and plant population were considered fixed effects. The row spacing, plant population, year, and their interaction means were subjected to a regression analysis using the software SigmaPlot 10.0. The criterion adopted to select a model was the magnitude of the regression coefficients (p < 0.05).
Results and discussion
The growing conditions, particularly the rainfall, were different between 2008 and 2009 (Figure 1). The rainfall during the castor cycle totaled 359 mm in 2008 and 655 mm in 2009; the 38-yr average is 555 mm. Because of the greater amount and better distribution of rainfall in 2009, the castor cycle was 36 days longer than in 2008. According to Souza et al. (2007), the excessive availability of water can favor the vegetative growth of castor plants and delay flowering and fruit ripening.
The row spacing, plant population, and their interaction were significant to the plant survival rate. Increasing the plant population density, particularly in a wide row spacing, decreased plant survival (Figure 2A), most likely because this combination led to higher plant densities within a row. The widest row spacing (0.90 m) combined with the highest plant population (70,000 plants ha-1) reduced the plant survival rate by approximately 11.4% compared to the combination of the narrowest row spacing (0.45 m) and sparsest plant population (25,000 plants ha-1) (Figure 2A). The plant population density had a greater effect on plant survival rate than the row spacing. Although weaker plants died before harvest because they were less competitive under a higher plant density, a lower plant density provided better conditions for proper growth. Soratto et al. (2012) also observed decreasing survival of castor plants due to an increasing number of plants per area (i.e., higher plant densities within a row). Similarly, Cox and Cherney (2011) found that higher plant densities in a row reduced the survival of soybean plants. In these situations, there is strong competition among the plants for water, light, and nutrients, and the weaker plants may not complete their growth cycle.

The basal stem diameter was more strongly affected by the plant population than by the row spacing (Figure 2B). Increasing the plant population reduced the stem diameter in all row spacings, but the stem diameter decreased more in the 0.90 m row spacing compared to the narrower row spacings. Distinct plant arrangements may affect the quality and quantity of the sunlight intercepted by plants. Kittock and Williams (1970) also found that an increased number of plants within a row reduced the diameter of castor stems. Bizinoto et al. (2010) found that the castor stem diameter decreased in response to an increased plant density. According to these authors, the reduction in stem diameter was possibly due to the etiolation caused by a high competition for light at a high plant density within a row or a dense plant population. Because castor plants with thin stems are favorable for mechanical harvesting (LOPES et al., 2008), our results indicate that the implementation of a high plant population can promotes plants with thinner stems, thus facilitating mechanical harvesting techniques (Figure 2B).
Plant height was not significantly affected by the row spacing and plant population, and the average plant height was 1.15 m. Kittock and Williams (1970) and Bizinoto et al. (2010) also found no effect of plant density on the height of castor plants. The effects of plant density on plant height is very complex and depends on both environmental factors and plant architecture. Plant growth is slow under the low water availability and temperature during the fall-winter cropping season, and the effect of row spacing and plant population on plant height appears to be lower than that observed in the spring-summer season, a time when the conditions are more favorable to plant growth (SEVERINO et al., 2006a and b; SORATTO et al., 2012).
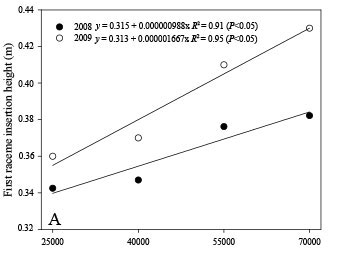
The height of insertion of the first raceme was linearly increased by increments in the plant population in both years, regardless of the row spacing (Figure 3A). In addition, the insertion height differences between the lower and higher plant populations were greater in the second year compared to the previous year, and more available water (Figure 1) may have caused stem etiolation. Kittock and Williams (1970), Bizinoto et al. (2010), and Soratto et al. (2012) reported that higher plant densities increased the first raceme insertion height of castor plants, without affecting plant height.
An increasing plant population decreased the number of racemes per plant at any row spacing (Figure 2C); similar results were found by Kittock and Williams (1970) and Bizinoto et al. (2010). The number of racemes per plant increased with increasing row spacing up to 0.65 m and then decreased with wider row spacings. The greatest number of racemes per plant was estimated for the sparsest plant population (25,000 plants ha-1) and the 0.65 m row spacing, i.e., plants more equidistantly distributed within (0.62 m) and between rows (0.65 m), with 1.62 plants m-1 within row. Whenever plants are more widely spaced, there is a larger area for light interception and lower competition or interference among them. Thus, photoassimilate production is higher, and plants display the appropriate architecture, which allows them to express their potential by forming more racemes (BIZINOTO et al., 2010; KITTOCK; WILLIAMS, 1970).
The number of seeds per raceme decreased linearly as the plant population increased in both years, regardless of row spacing (Figure 3B). It was notable that the number of seeds per raceme obtained in 2008 was higher than in 2009, particularly for the lower plant populations, possibly because of the higher rainfall in the second year (Figure 1), which occurred at the fruit formation stage (June and July). A high relative humidity increases the incidence of gray mold in racemes and, thus, may reduce the number of seeds per raceme (SUSSEL et al., 2009).
The seed weight was not significantly affected by the factors studied. The seed weight average was higher in 2009 (463 mg seed-1) than in 2008 (376 mg seed-1), possibly because of the increased water availability during the seed formation and filling stages (Figure 1) and the lower number of seeds per raceme (Figure 3B).
Seed yield was affected by the row spacing × plant population interaction, and both row spacing and plant population had quadratic effects on the grain yield (Figure 4A). Considering the average for the two years, the highest seed yield (2,248 kg ha-1) was estimated for the combination of 0.64 m row spacing and 50,107 plants ha-1 (i.e., 3.2 plants m-1). However, the effects of plant population and row spacing on the seed yield were not as strong as observed for the number of racemes per plant and seeds per raceme (Figure 2C and 3B). No correlation between this yield components and the seed yield was observed, which indicates that the castor plants compensated at the lower population densities while increasing the number of racemes per plant and seeds per raceme, without an equivalent variation in the seed yield. Severino et al. (2006a and b) and Carvalho et al. (2010) found higher castor seed yields by increasing the plant population, but those studies did not include very high plant populations, such as those used in the present research. Soratto et al. (2012) reported that, for a high plant population (55,000 plants ha-1), the seed yield of two castor genotypes in fall-winter was greater at more equidistant plant arrangements because the plant density decreased within a row, promoting a better plant distribution in the area and lower competition among the plants.
The oil content in the castor seeds was not significantly affected by the treatments. Averaging the two years, the seed oil content of the subplot samples varied within the range of 43.4% to 48.5%, values that are similar to those reported in the literature (SEVERINO et al., 2006a and b). According to Severino et al. (2006b), the oil content in castor seeds is a characteristic of high hereditability. Kittock and Williams (1970) also observed no influence of the plant population on oil content. According to Koutroubas et al. (1999), higher seed yields are usually obtained at the expense of the oil content, and there is a negative correlation between these two variables; however, this correlation was not found in the present study.
Because the seed oil content was not influenced by the treatments, the seed yield and oil yield were similarly influenced by the row spacing and plant population (Figure 4A and B). The seed yield was positively and strongly correlated to the oil yield (r = 0.97, p < 0.001) (i.e., the oil yield was mainly determined by the seed yield).
The results of this study showed that the castor plant responses to changes in the plant population and arrangement are very complex and may be dependent on the weather conditions. At a plant population that is higher than generally recommended and used in Brazil (CARVALHO et al., 2010; SEVERINO et al., 2006a and b), the castor plants compensated for a lower plant population with a high number of racemes per plant and seeds per raceme in such a way that confined the seed and oil yields to a narrow range. Thus, for castor genotype FCA-PB grown in the fall-winter season, higher plant populations (~50,000 plants ha-1) and narrower row spacing (~0.65 m) can be useful for providing greater seed and oil yields (Figure 4A and B) and plants with thinner stems (Figure 2B), thus facilitating mechanical harvesting.
Conclusion
A high plant density within a row caused a reduction in the survival rate of castor plants. The first raceme insertion height was increased by increasing the plant population, particularly in the year with the higher rainfall. The castor plants compensated for a low plant population with a high number of racemes per plant and seeds per raceme such that the seed and oil yields varied only within a narrow range. The maximum seed yield of castor genotype FCA-PB in the fall-winter season was achieved with an estimated 0.64 m row spacing and 50,107 plants ha-1 (i.e., 3.2 plants m-1).
Acknowledgements
To FAPESP (State of São Paulo Research Foundation) for supporting this research and providing scholarship to the first author (Proc. 2007/05851-8). To CNPq (The National Council for Scientific and Technological Development) for providing research award to the second author. To Dr. Luiz Alberto Colnago, from Embrapa Agricultural Instrumentation, for assistance in determining seed oil content.
References
BABITA, M.; MAHESWARI, M.; RAO, L. M.; SHANKER, A. K.; RAO, D. G. Osmotic adjustment, drought tolerance and yield of castor (Ricinus communis L.) hybrids. Environmental and Experimental Botany, v. 69, n. 3, p. 243-249, 2010.
BALDWIN, B. S.; COSSAR, R. D. Castor yield in response to planting date at four locations in the south-central United States. Industrial Crops and Products, v. 29, n. 2-3, p. 316-319, 2009.
BARNES, D. J.; BALDWIN, B. S.; BRAASCH, D. A. Degradation of ricin in castor seed meal by temperature and chemical treatment. Industrial Crops and Products, v. 29, n. 2-3, p. 509-515, 2009.
BIZINOTO, T. K. M. C.; OLIVEIRA; E. G.; MARTINS; S. B.; SOUZA, S. A.; GOTARDO, M. Cultivo da mamoneira influenciada por diferentes populações de plantas. Bragantia, v. 69, n. 2, p. 367-370, 2010.
CARVALHO, E. V.; SÁ, C. H. A. C.; COSTA, J. L.; AFFÉRRI, F. S.; SIEBENEICHLER, S. C. Densidade de plantio em duas cultivares de mamona no Sul do Tocantins. Revista Ciência Agronômica, v. 41, n. 3, p. 387-392, 2010.
COLNAGO, L. A.; ENGELSBERG, M.; SOUZA, A. A.; BARBOSA, L. L. High-throughput, non-destructive determination of oil content in intact seeds by continuous wave-free precession NMR. Analytical Chemistry, v. 79, n. 3, p. 1271-1274, 2007.
COX, W. J.; CHERNEY, J. H. Growth and yield responses of soybean to row spacing and seeding rate. Agronomy Journal, v. 3, n. 1, p. 123-128, 2011.
KITTOCK, D. L.; WILLIAMS, J. H. Effects of plant population on castorbean yield. Agronomy Journal, v. 62, n. 4, p. 527-529, 1970.
KOUTROUBAS, S. D.; PAPAKOSTA, D. K.; DOITSINIS, A. Adaptation and yielding ability of castor plant (Ricinus communis L.) genotypes in a Mediterranean climate. European Journal of Agronomy, v. 11, n. 3-4, p. 227-237, 1999.
LOPES, F. F. M.; BELTRÃO, N. E. M.; LOPES NETO, J. P.; PEDROZA, J. P. Crescimento inicial de genótipos de mamoneira com sementes submetidas ao envelhecimento acelerado. Revista Brasileira de Oleaginosas e Fibrosas, v. 12, n. 1, p. 69-79, 2008.
SAVY FILHO, A. Mamona: tecnologia agrícola. Campinas: Emopi, 2005.
SEVERINO, L. S.; AULD, D. L.; BALDANZI, M.; CÂNDIDO, M. J. D.; CHEN, G.; CROSBY, W.; TAN, D.; HE, X.; LAKSHMAMMA, P.; LAVANYA, C.; MACHADO, O. L. T.; MIELKE, T.; MILLER, T. D.; MORRIS, J. B.; MORSE, S. A.; NAVAS, A. A.; SOARES, D. J.; SOFIATTI, V.; WANG, M. L.; ZANOTTO, M. D.; ZIELER, H. A Review on the challenges for increased production of castor. Agronomy Journal, v. 104, n. 4, p. 853-880, 2012.
SEVERINO, L. S.; COELHO, D. K.; MORAES, C. R. A.; GONDIM, T. M. S.; VALE, L. S. Otimização do espaçamento de plantio para a mamoneira cultivar BRS Nordestina. Revista Brasileira de Oleaginosas e Fibrosas, v. 10, n. 1-2, p. 993-999, 2006a.
SEVERINO, L. S.; MORAES, C. R. A.; GONDIM, T. M. S.; CARDOSO, G. D.; BELTRÃO, N. E. M. Crescimento e produtividade da mamoneira influenciada por plantio em diferentes espaçamentos entre linhas. Revista Ciência Agronômica, v. 37, n. 1, p. 50-54, 2006b.
SILVA, A. G.; CRUSCIOL, C. A. C.; SORATTO, R. P.; COSTA, C. H. M.; FERRARI NETO, J. Produção de fitomassa e acúmulo de nutrientes por plantas de cobertura e cultivo da mamona em sucessão no sistema plantio direto. Ciência Rural, v. 40, n. 10, p. 2092-2098, 2010.
SORATTO, R. P.; SOUZA-SCHLICK, G. D.; FERNANDES, A. M.; ZANOTTO, M. D.; CRUSCIOL, C. A. C. Narrow row spacing and high plant population to short height castor genotypes in two cropping seasons. Industrial Crops and Products, v. 35, n. 1, p. 244-249, 2012.
SOUZA, A. S.; TÁVORA, F. J. A. F.; PITOMBEIRA, J. B.; BEZERRA, F. M. L. Épocas de plantio e manejo da irrigação para a mamoneira. II - crescimento e produtividade. Revista Ciência Agronômica, v. 38, n. 4, p. 422-429, 2007.
SOUZA-SCHLICK, G. D.; SORATTO, R. P.; PASQUALI, C. B.; FERNANDES, A. M. Desempenho da mamoneira IAC 2028 em função do espaçamento entre fileiras e população de plantas na safrinha. Bragantia, v. 70, n. 3, p. 519-528, 2011.
SUSSEL, A. A. B.; POZZA, E. A.; CASTRO, H. A. Elaboração e validação de escala diagramática para avaliação da severidade do mofo cinzento em mamoneira. Tropical Plant Pathology, v. 34, n. 3, p. 186-191, 2009.
ZHOU, G.; MA, B. L.; LI, J.: FENG, C.; LU, J.; QIN, P. Determining salinity threshold level for castor bean emergence and stand establishment. Crop Science, v. 50, n. 5, p. 2030-2036, 2010.
Received on June 3, 2012
Accepted on October 31, 2012
License information: This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
- BABITA, M.; MAHESWARI, M.; RAO, L. M.; SHANKER, A. K.; RAO, D. G. Osmotic adjustment, drought tolerance and yield of castor (Ricinus communis L.) hybrids. Environmental and Experimental Botany, v. 69, n. 3, p. 243-249, 2010.
- BALDWIN, B. S.; COSSAR, R. D. Castor yield in response to planting date at four locations in the south-central United States. Industrial Crops and Products, v. 29, n. 2-3, p. 316-319, 2009.
- BARNES, D. J.; BALDWIN, B. S.; BRAASCH, D. A. Degradation of ricin in castor seed meal by temperature and chemical treatment. Industrial Crops and Products, v. 29, n. 2-3, p. 509-515, 2009.
- BIZINOTO, T. K. M. C.; OLIVEIRA; E. G.; MARTINS; S. B.; SOUZA, S. A.; GOTARDO, M. Cultivo da mamoneira influenciada por diferentes populações de plantas. Bragantia, v. 69, n. 2, p. 367-370, 2010.
- CARVALHO, E. V.; SÁ, C. H. A. C.; COSTA, J. L.; AFFÉRRI, F. S.; SIEBENEICHLER, S. C. Densidade de plantio em duas cultivares de mamona no Sul do Tocantins. Revista Ciência Agronômica, v. 41, n. 3, p. 387-392, 2010.
- COLNAGO, L. A.; ENGELSBERG, M.; SOUZA, A. A.; BARBOSA, L. L. High-throughput, non-destructive determination of oil content in intact seeds by continuous wave-free precession NMR. Analytical Chemistry, v. 79, n. 3, p. 1271-1274, 2007.
- COX, W. J.; CHERNEY, J. H. Growth and yield responses of soybean to row spacing and seeding rate. Agronomy Journal, v. 3, n. 1, p. 123-128, 2011.
- KITTOCK, D. L.; WILLIAMS, J. H. Effects of plant population on castorbean yield. Agronomy Journal, v. 62, n. 4, p. 527-529, 1970.
- KOUTROUBAS, S. D.; PAPAKOSTA, D. K.; DOITSINIS, A. Adaptation and yielding ability of castor plant (Ricinus communis L.) genotypes in a Mediterranean climate. European Journal of Agronomy, v. 11, n. 3-4, p. 227-237, 1999.
- LOPES, F. F. M.; BELTRÃO, N. E. M.; LOPES NETO, J. P.; PEDROZA, J. P. Crescimento inicial de genótipos de mamoneira com sementes submetidas ao envelhecimento acelerado. Revista Brasileira de Oleaginosas e Fibrosas, v. 12, n. 1, p. 69-79, 2008.
- SAVY FILHO, A. Mamona: tecnologia agrícola. Campinas: Emopi, 2005.
- SEVERINO, L. S.; AULD, D. L.; BALDANZI, M.; CÂNDIDO, M. J. D.; CHEN, G.; CROSBY, W.; TAN, D.; HE, X.; LAKSHMAMMA, P.; LAVANYA, C.; MACHADO, O. L. T.; MIELKE, T.; MILLER, T. D.; MORRIS, J. B.; MORSE, S. A.; NAVAS, A. A.; SOARES, D. J.; SOFIATTI, V.; WANG, M. L.; ZANOTTO, M. D.; ZIELER, H. A Review on the challenges for increased production of castor. Agronomy Journal, v. 104, n. 4, p. 853-880, 2012.
- SEVERINO, L. S.; COELHO, D. K.; MORAES, C. R. A.; GONDIM, T. M. S.; VALE, L. S. Otimização do espaçamento de plantio para a mamoneira cultivar BRS Nordestina. Revista Brasileira de Oleaginosas e Fibrosas, v. 10, n. 1-2, p. 993-999, 2006a.
- SEVERINO, L. S.; MORAES, C. R. A.; GONDIM, T. M. S.; CARDOSO, G. D.; BELTRÃO, N. E. M. Crescimento e produtividade da mamoneira influenciada por plantio em diferentes espaçamentos entre linhas. Revista Ciência Agronômica, v. 37, n. 1, p. 50-54, 2006b.
- SILVA, A. G.; CRUSCIOL, C. A. C.; SORATTO, R. P.; COSTA, C. H. M.; FERRARI NETO, J. Produção de fitomassa e acúmulo de nutrientes por plantas de cobertura e cultivo da mamona em sucessão no sistema plantio direto. Ciência Rural, v. 40, n. 10, p. 2092-2098, 2010.
- SORATTO, R. P.; SOUZA-SCHLICK, G. D.; FERNANDES, A. M.; ZANOTTO, M. D.; CRUSCIOL, C. A. C. Narrow row spacing and high plant population to short height castor genotypes in two cropping seasons. Industrial Crops and Products, v. 35, n. 1, p. 244-249, 2012.
- SOUZA, A. S.; TÁVORA, F. J. A. F.; PITOMBEIRA, J. B.; BEZERRA, F. M. L. Épocas de plantio e manejo da irrigação para a mamoneira. II - crescimento e produtividade. Revista Ciência Agronômica, v. 38, n. 4, p. 422-429, 2007.
- SOUZA-SCHLICK, G. D.; SORATTO, R. P.; PASQUALI, C. B.; FERNANDES, A. M. Desempenho da mamoneira IAC 2028 em função do espaçamento entre fileiras e população de plantas na safrinha. Bragantia, v. 70, n. 3, p. 519-528, 2011.
- SUSSEL, A. A. B.; POZZA, E. A.; CASTRO, H. A. Elaboração e validação de escala diagramática para avaliação da severidade do mofo cinzento em mamoneira. Tropical Plant Pathology, v. 34, n. 3, p. 186-191, 2009.
- ZHOU, G.; MA, B. L.; LI, J.: FENG, C.; LU, J.; QIN, P. Determining salinity threshold level for castor bean emergence and stand establishment. Crop Science, v. 50, n. 5, p. 2030-2036, 2010.
Publication Dates
-
Publication in this collection
07 Oct 2014 -
Date of issue
Dec 2014
History
-
Received
03 June 2012 -
Accepted
31 Oct 2012






