Abstract
Abstract: Insects are the most diverse group of animals. They can be infected by an extraordinary diversity of viruses. Among them, arthropod-borne viruses (arboviruses) can be transmitted to humans. High-throughput sequencing of small RNAs from insects provides insight into their virome, which may help understand the dynamics of vector borne infectious diseases. Furthermore, investigating the mechanisms that restrict viral infections in insects points to genetic innovations that may inspire novel antiviral strategies.
Key words
antiviral immunity; insects; metagenomics; restriction factors; RNA interference; virome
DIVERSITY OF INSECTS AND VIRUSES
With around 1 million known species, insects are the largest group of multicellular organisms, representing over 70% of animal species (Stork 2018, Mora et al. 2011). With an origin dating back to the Early Ordovician (~480 million years ago (Mya)), ancestors of insects were among the first animals to colonize terrestrial and freshwater ecosystems and have undergone major expansions, culminating in the spectacular diversification of holometabolous insects (Hymenoptera, Coleoptera, Diptera, Lepidoptera) during Early Cretaceous (~120Mya) (Misof et al. 2014). Insects can be credited with major innovations, such as flight and establishment of social groups. Importantly, they have undoubtedly contributed to shape the planet’s biota and actively interact with other multicellular Eukaryotes such as plants and vertebrates. Like them, diverse insects are exposed to a large panel of infectious microorganisms (Figure 1), which they control through their innate immune system.

Diversity of insects and viruses. A plethora of known insect viruses and insect hosts is depicted on the left and right panels, respectively. Insects broadly evolved to resist infection, so investigating the evolutionarily conserved antiviral strategies in these animals can shed light on new host factors and restriction factors that affect viral infection. The pie chart represents the percentage of D. melanogaster genes that are conserved or non-conserved in the human genome. D. melanogaster exclusive genes are highlighted among the non-conserved genes. Adapted from Martins et al. 2016.
Among infectious microbes, viruses represent a particular threat because they offer few intrinsic targets for inhibition by antiviral molecules. This is because they consist in their simplest form in a nucleic acid encapsulated in a protein shell, and hijack molecular machineries from host cells to complete their replication cycle. Therefore, viruses exert great selective pressure on their host to evolve resistance pathways. These, in turn, favor the adaptation of viruses to escape antiviral mechanisms. This arms race results in the diversification of host-defense and virus escape mechanisms. As a result, it can be highly instructive to investigate virus–host interactions in a range of animals, to sample in depth the diversity of antiviral strategies. In light of their aforementioned diversity, insects represent an interesting group of animals for comparative studies (Martins et al. 2016, Marques and Imler 2016). A further reason to investigate insect-virus interactions is that hematophagous insects, for example Aedes mosquitoes, are vectors of medically important viral diseases such as Zika, Dengue, Chikungunya and Yellow fever.
The recent advances in high-throughput sequencing technologies have opened the way to the characterization of the virome (i.e. the genetic diversity of viruses in a biological sample). In a landmark article, Yong-Zhen Zhang and colleagues analyzed the transcriptome of more than 220 invertebrate species covering nine animal phyla and reported the identification of close to 1,500 new viruses (Shi et al. 2016). Thus, infection by one or several viruses is common in invertebrates. The genetic diversity of these viruses surpassed that described previously (Figure 1). While many of them could not be incorporated into the current virus classification, newly identified viruses fell between families and genera, thus filling major phylogenetic gaps and revealing that viruses form a more continual spectrum of phylogenetic diversity (Shi et al. 2016). A more detailed analysis focusing on 70 arthropod species and negative-sense RNA viruses, which include important pathogens causing a variety of diseases in humans (flu, rabies, encephalitis, hemorrhagic fever), led to the discovery of 112 new viruses. It revealed that arthropods are likely to have participated in the evolution of these important viruses (Dudas and Obbard 2015, Li et al. 2015). Indeed, much of the diversity of negative-sense RNA viruses found in plants and vertebrate animals falls within the genetic diversity of viruses associated with arthropods. Arthropods and insects in particular can live in large and dense populations, facilitating propagation and transmission of viruses. In addition, the well described close interaction between insects and plants supports the hypothesis that negative-sense RNA viruses, including vertebrate-specific ones, derived from arthropod dependent viruses (Li et al. 2015).
KEEPING A WATCH ON THE VIROME OF INSECT VECTORS
The number and diversity of viruses identified in insect samples may have public health significance since mosquitoes and other insect species can transmit diseases to humans. Indeed, these viruses may include putative emerging pathogens, or may affect the dynamics of transmission of known viral pathogens such as Dengue, Zika or Chikungunya viruses. While large scale sequencing of the transcriptome is commonly used to identify viruses (e.g. Li et al. 2015, Shi et al. 2016), it suffers from limitations such as dilution among cellular RNAs, which may result in a majority of non-viral sequences in the library, and confusion between transcripts derived from endogenous viral elements (EVEs) and replicating viruses. Another major limitation is that identification of viruses relies on sequence similarity comparisons to reference databases. Indeed, a significant number of novel sequences identified in large scale sequencing studies remain of unknown origin because they do not bear similarity to known sequences. These unassigned sequences are sometimes referred to as the “dark matter” of metagenomics (Oh et al. 2014).
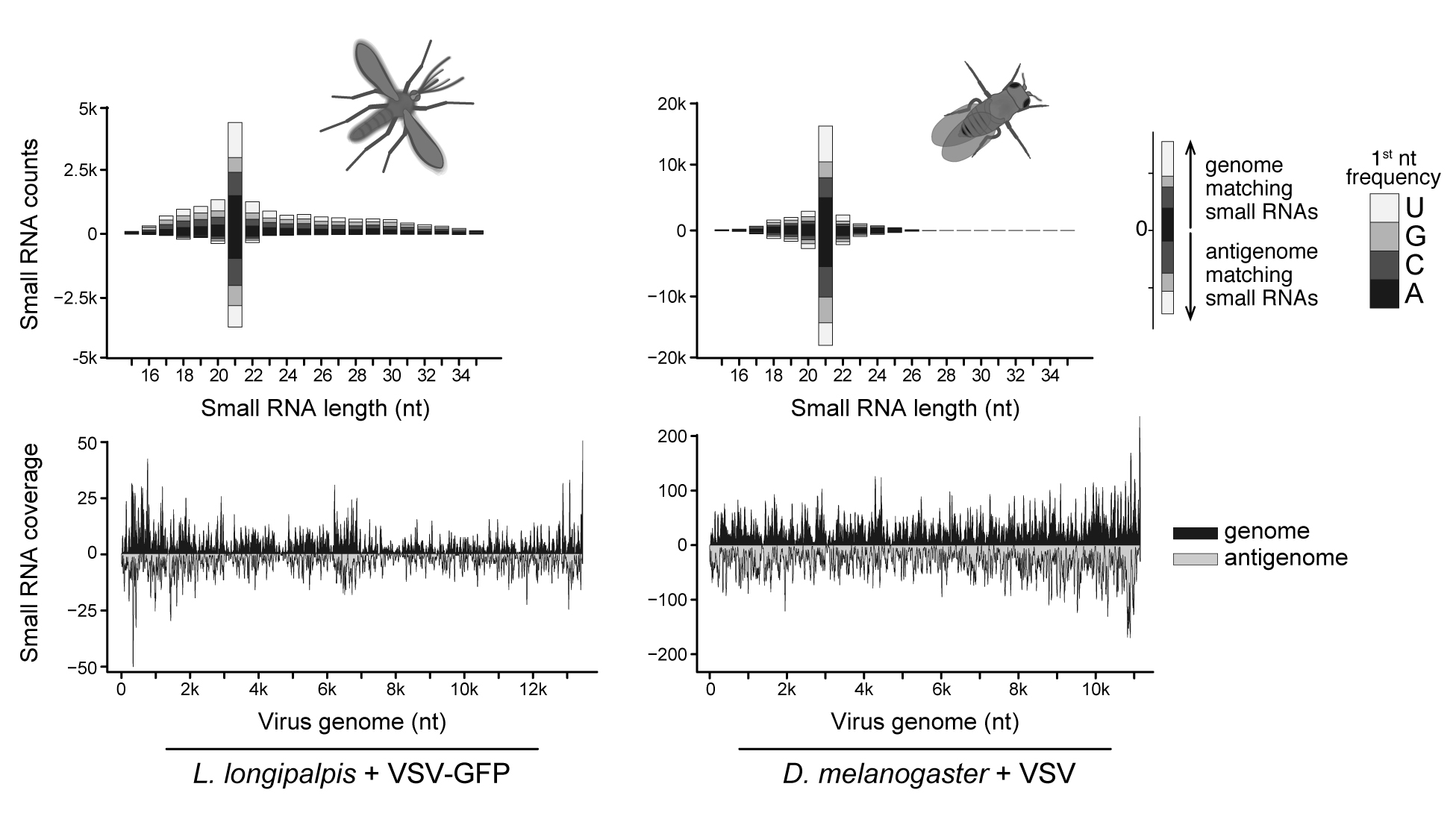
RNA interference (RNAi) represents a major facet of antiviral innate immunity in insects, which can be exploited to explore the virome of insects. The major antiviral RNAi pathway is the small interfering (si) RNA pathway. It involves the RNaseIII enzyme Dicer-2, which senses replicating viral RNAs and processes them into 21 nucleotide (nt)-long siRNAs (reviewed in Paro et al. 2015, Aguiar et al. 2016). These siRNAs originate from both the genome and the antigenome of the virus, as observed during infection by the rhabdovirus Vesicular Stomatitis virus (VSV) in the model organism Drosophila melanogaster and the sandfly Lutzomyia longipalpis, which is one natural vector for this arthropod-borne virus (Figure 2) (Mueller et al. 2010, Marques et al. 2013, Ferreira et al. 2018). In Aedes mosquitoes, another RNAi pathway called piRNA pathway, which involves 24-29nt long Piwi-interacting (pi) RNAs, is also activated upon viral infections (Morazzani et al. 2012, Aguiar et al. 2015, Dietrich et al. 2017). Viral contigs can be assembled from large scale sequencing of small RNAs and this strategy has been used for virus identification in insects and plants, which also rely on RNAi for antiviral immunity (Wu et al. 2010, Kreuze et al. 2009). We have conducted a comparative analysis of long and small RNA sequencing for virus identification and showed that viral sequences are enriched in the small RNA fraction (Aguiar et al. 2015). Furthermore, the average size of viral contigs assembled from small RNA libraries was longer, resulting in larger coverage of viral genomes than the traditional long RNA libraries. Hence, sequencing of virus-derived small RNAs produced by the immune response of the insect host represents a powerful strategy to identify viral sequences.

Vesicular Stomatitis virus (VSV)-derived siRNAs in D. melanogaster and L. longipalpis. Activation of the siRNA pathway is a conserved response to virus infection in insects. Virus-derived siRNAs are mostly 21nt long small RNAs and show symmetrical base preference and polarity. siRNAs originate from both the viral genome and antigenome and can be detected along the entire extension of the virus sequence. Adapted from Ferreira et al. 2018 and Mueller et al. 2010.
While applying this strategy to three Dipteran insect species (D. melanogaster, Ae. aegypti, L. longipalpis), we observed that the size profile of virus-derived small RNAs was unique for many viruses (Figure 3). These differences include production of virus derived piRNAs for some viruses (e.g. Phasi Charoen like-virus (PCLV) but not Humaita-Tubiacanga virus (HTV) in Ae. aegypti), virus-derived small RNAs smaller than 21nt, possibly resulting from degradation (e.g. Drosophila uncharacterized virus (DUV) in D. melanogaster) or lack of symmetry between (+) and (-) strands of viral RNA (e.g. Lutzomyia Piaui nodavirus (LPNV) in L. longipalpis). Based on these observations, we used a Z-score to normalize the small RNA size profiles and generated heatmaps for each contig that could be subjected to hierarchical clustering based on pairwise correlations (Aguiar et al. 2015). This allowed us to evaluate the relationship between contigs and to assign a viral origin to contigs that lack significant sequence similarity to reference databases.

Profiles of virus-derived small RNAs in dipteran insects. Viral RNA can be recognized by different antiviral surveillance mechanisms, thus generating specific virus-derived small RNA profiles during infection. These distinct patterns can be analyzed and provide extensive information about their source and biogenesis, such as the activation of siRNA (e.g. PCLV, HTV, DRV and DUV) and piRNA (e.g. PCLV) pathways. Even a virus-derived small RNA profile consistent with RNA degradation (e.g. Lutzomyia Piaui reovirus 1 (LPRV1), Lutzomyia Piaui reovirus 2 (LPRV2) and LPNV) can be indicative that the virus escapes RNAi during infection. Adapted from Aguiar et al. 2015.
INSECTS AS A RESERVOIR OF INNOVATIONS FOR ANTIVIRAL MECHANISMS
Besides RNAi, insects also rely on induced responses and restriction factors to control viral infections. The evolutionarily conserved Toll and IMD pathways, which regulate transcription factors of the NF-κB family and were initially characterized for their role in antifungal and antibacterial immunity, have been proposed to play a role in antiviral immunity in drosophila (reviewed in Mussabekova et al. 2017). More recently, a new pathway, involving the ortholog of the factor STING (STimulator of INterferon Genes), the kinase IKKβ and the NF-κB transcription factor Relish was shown to control infection by picorna-like viruses in drosophila (Goto et al. 2018). Of note, no dSTING ortholog is found in mosquito genomes, suggesting that different mechanisms are elicited in response to infection in these insects. The cytokine-induced JAK/STAT pathway also participates in antiviral immunity in flies and mosquitoes (Dostert et al. 2005, Souza-Neto et al. 2009, Paradkar et al. 2012). The genes upregulated by these pathways in response to viral infections encode components of signaling pathways (e.g. dSTING), cytokines but also antiviral factors, which for the most part remain to be characterized. In addition, viruses are also controlled by constitutively expressed restriction factors (Mussabekova et al. 2017). Some of the inducible antiviral factors and restriction factors are evolutionarily conserved and the functions of their mammalian homologues point to mechanisms for virus inhibition (e.g. ref(2)P/p62 and autophagy, Carré-Mlouka et al. 2007; heat shock proteins and protein folding, Merkling et al. 2015). Interestingly, others are not conserved and represent insect innovations.
Two drosophila genes, pastrel and nazo, are examples of such innovations. Both are involved in the restriction of picorna-like viruses in drosophila. A cluster of polymorphisms in and around the D. melanogaster gene pastrel is associated with resistance or susceptibility to the Drosophila C virus (DCV) (Cao et al. 2017). Silencing of pastrel by RNAi results in increased DCV replication, revealing that Pastrel serves as a restriction factor. Orthologues of pastrel can be found in the genomes of drosophilids and a subset of Diptera, although it is not known yet if they serve in antiviral host defense in these species as well. The expression of nazo is upregulated by the STING/IKKβ pathway in response to DCV infection (Goto et al. 2018). Silencing of nazo results in increased DCV infection, whereas its ectopic expression in tissue culture cells efficiently suppresses replication of the virus. Orthologues of Nazo are present in all animals, where their function is unknown. The gene has been duplicated in D. melanogaster and a subset of drosophilids, and only Nazo has antiviral activity suggesting that a yet uncharacterized evolutionarily conserved cellular factor has been recruited in fruit flies to acquire a novel antiviral function (Goto et al. 2018). Of note, both Pastrel and Nazo also control replication of Cricket Paralysis virus, which belongs to the same family as DCV (Dicistroviridae). This indicates that the two drosophila factors target conserved steps of the replication cycle of these viruses and that the characterization of their mode of action will point to one or more weak spots in Dicistroviridae. This knowledge may be exploited to develop novel antiviral agents targeting the related Picornaviridae, which encompass several important human pathogens such as poliovirus and Hepatitis A virus.
CONCLUSIONS
The diversity of insect hosts and of the viruses infecting them provides an unparalleled opportunity to decipher (i) the repertoire of antiviral strategies in animals, and (ii) the suppressor mechanisms evolved in viruses as countermeasures. Insects, with their fantastic diversity, represent a reservoir of unique antiviral strategies, which could be inspirational for the design of new therapeutic agents. In the specific case of vector mosquitoes, characterization of the virome in wild populations is an important step to uncover persistent infections by viruses expressing suppressors of the immune response that may affect the dynamics of transmission of other viruses such as Dengue, Chikungunya or Zika.
ACKNOWLEGMENTS
This work was funded by the University of Strasbourg Institute for Advanced Studies, CNRS, the Institut Universitaire de France, the Programme Investissements d’Avenir (ANR-10-LABX-0036 and ANR-11-EQPX-0022), the Agence Nationale de la Recherche (ANR-17-CE15- 0014-01).
REFERENCES
- AGUIAR ER, OLMO RP and MARQUES JT. 2016. Virus-derived small RNAs: molecular footprints of host-pathogen interactions. Wiley Interdiscip Rev RNA 7: 824-837.
- AGUIAR ER et al. 2015. Sequence-independent characterization of viruses based on the pattern of viral small RNAs produced by the host. Nucleic Acids Res 43: 6191-6206.
- CAO C, COGNI R, BARBIER V and JIGGINS FM. 2017. Complex Coding and Regulatory Polymorphisms in a Restriction Factor Determine the Susceptibility of Drosophila to viral infection. Genetics 206: 2159-2173.
- CARRÉ-MLOUKA A, GAUMER S, GAY P, PETITJEAN AM, COULONDRE C, DRU P, BRAS F, DEZÉLÉE S and CONTAMINE D. 2007. Control of sigma virus multiplication by the ref(2)P gene of Drosophila melanogaster: an in vivo study of the PB1 domain of Ref(2)P. Genetics 176: 409-419.
- DIETRICH I et al. 2017. RNA Interference Restricts Rift Valley Fever Virus in Multiple Insect Systems. mSphere 2: e00090-17.
- DOSTERT C, JOUANGUY E, IRVING P, TROXLER L, GALIANA-ARNOUX D, HETRU C, HOFFMANN JA and IMLER JL. 2005. The Jak-STAT signaling pathway is required but not sufficient for the antiviral response of drosophila. Nat Immunol 6: 946-953.
- DUDAS G and OBBARD DJ. 2015. Are arthropods at the heart of virus evolution? Elife 4: e06837.
- FERREIRA FV, AGUIAR ERGR, OLMO RP, DE OLIVEIRA KPV, SILVA EG, SANT’ANNA MRV, GONTIJO NF, KROON EG, IMLER JL and MARQUES JT. 2018. The small non-coding RNA response to virus infection in the Leishmania vector Lutzomyia longipalpis. PLoS Negl Trop Dis 12: e0006569.
- GOTO A et al. 2018. The Kinase IKKbeta Regulates a STING- and NF-kappaB-Dependent Antiviral Response Pathway in Drosophila. Immunity 49(2): 225-234.e224.
- KREUZE JF, PEREZ A, UNTIVEROS M, QUISPE D, FUENTES S, BARKER I and SIMON R. 2009. Complete viral genome sequence and discovery of novel viruses by deep sequencing of small RNAs: a generic method for diagnosis, discovery and sequencing of viruses. Virology 388: 1-7.
- LI CX, SHI M, TIAN JH, LIN XD, KANG YJ, CHEN LJ, QIN XC, XU J, HOLMES EC and ZHANG YZ. 2015. Unprecedented genomic diversity of RNA viruses in arthropods reveals the ancestry of negative-sense RNA viruses. Elife 4: e05378.
- MARQUES JT and IMLER JL. 2016. The diversity of insect antiviral immunity: insights from viruses. Curr Opin Microbiol 32: 71-76.
- MARQUES JT, WANG JP, WANG X, DE OLIVEIRA KP, GAO C, AGUIAR ER, JAFARI N and CARTHEW RW. 2013. Functional specialization of the small interfering RNA pathway in response to virus infection. PLoS Pathog 9: e1003579.
- MARTINS N, IMLER JL and MEIGNIN C. 2016. Discovery of novel targets for antivirals: learning from flies. Curr Opin Virol 20: 64-70.
- MERKLING SH, OVERHEUL GJ, VAN MIERLO JT, ARENDS D, GILISSEN C and VAN RIJ RP. 2015. The heat shock response restricts virus infection in Drosophila. Sci Rep 5: 12758.
- MISOF B et al. 2014. Phylogenomics resolves the timing and pattern of insect evolution. Science 346: 763-767.
- MORA C, TITTENSOR DP, ALD S, SIMPSON AGB and WORM B. 2011. How many species are there on Earth and in the ocean? PLoS biology 9: e1001127.
- MORAZZANI EM, WILEY MR, MURREDDU MG, ADELMAN ZN and MYLES KM. 2012. Production of virus-derived ping-pong-dependent piRNA-like small RNAs in the mosquito soma. PLoS pathogens 8: e1002470.
- MUELLER S, GAUSSON V, VODOVAR N, DEDDOUCHE S, TROXLER L, PEROT J, PFEFFER S, HOFFMANN JA, SALEH MC and IMLER JL. 2010. RNAi-mediated immunity provides strong protection against the negative-strand RNA vesicular stomatitis virus in Drosophila. Proc Natl Acad Sci U S A 107: 19390-19395.
- MUSSABEKOVA A, DAEFFLER L and IMLER JL. 2017. Innate and intrinsic antiviral immunity in Drosophila. Cell Mol Life Sci 74: 2039-2054.
- OH J, BYRD AL, DEMING C, CONLAN S, KONG HH, SEGRE JA and PROGRAM NCS. 2014. Biogeography and individuality shape function in the human skin metagenome. Nature 514: 59-64.
- PARADKAR PN, TRINIDAD L, VOYSEY R, DUCHEMIN JB and WALKER PJ. 2012. Secreted Vago restricts West Nile virus infection in Culex mosquito cells by activating the Jak-STAT pathway. Proc Natl Acad Sci U S A 109: 18915-18920.
- PARO S, IMLER JL and MEIGNIN C. 2015. Sensing viral RNAs by Dicer/RIG-I like ATPases across species. Curr Opin Immunol 32: 106-113.
- SHI M et al. 2016. Redefining the invertebrate RNA virosphere. Nature 540: 539-543.
- SOUZA-NETO JA, SIM S and DIMOPOULOS G. 2009. An evolutionary conserved function of the JAK-STAT pathway in anti-dengue defense. Proc Natl Acad Sci U S A 106: 17841-17846.
- STORK NE. 2018. How Many Species of Insects and Other Terrestrial Arthropods Are There on Earth? Annu Rev Entomol 63: 31-45.
- WU Q, LUO Y, LU R, LAU N, LAI EC, LI WX and DING SW. 2010. Virus discovery by deep sequencing and assembly of virus-derived small silencing RNAs. Proc Natl Acad Sci U S A 107: 1606-1611.