Resumos
Para verificar o efeito do estresse calórico (EC) nas concentrações plasmáticas de testosterona, triiodotironina (T3) e tiroxina (T4), oito bodes, das raças Saanen (n=4) e Alpina (n=4), foram mantidos em câmara bioclimática, sob condições de termoneutralidade (13,0ºC a 26,7ºC) durante 30 dias e, após um período (60 dias) de descanso, submetidos ao EC (23,7ºC a 34,0ºC) por 30 dias. Para minimizar as variações sazonais nos perfis hormonais devido ao fotoperíodo, durante toda fase experimental, incluindo a de adaptação em condições de termoneutralidade (30 dias), o fotoperíodo foi controlado utilizando-se alternância de dias longos (16h de luz e 8h de escuro) e de dias curtos (8h de luz e 16h de escuro) a cada 30 dias. As amostras de sangue foram coletadas duas vezes por semana durante cinco semanas. No conjunto das raças, o EC não influenciou (P>0,05) as concentrações de testosterona (1,8±0,2 vs 1,3±0,2ng/ml) e nem a de T4 (52,7±2,8 vs 50,0±2,8ng/ml). Houve declínio (P<0,01) das concentrações de T3 nos animais submetidos ao experimento (1,3±0,1 vs 1,0±0,1ng/ml), mas a redução foi observada somente nos bodes Saanen. Em ambas as raças, as concentrações de T3 e T4 variaram (P<0,01) conforme o dia da coleta das amostras de sangue. O EC foi suficiente para produzir uma resposta fisiológica com redução das concentrações plasmáticas de T3 em bodes das raças Saanen, mas não da raça Alpina, assim como não foi capaz de alterar os níveis plasmáticos de testosterona e nem de T4.
bode; estresse calórico; hormônios tireoidianos; testosterona
To verify the effect of heat stress (HS) on plasma testosterone, triiodothyronine (T3), and thyroxine (T4) concentrations, eight Saanen (n=4) and Alpine Brown (n=4) bucks were kept in climate chamber under thermal neutral conditions (13.0ºC to 26.7ºC) for 30 days. After a resting period (60 days), the same bucks were submitted to heat stress (23.7ºC to 34.0ºC) for another 30 days. To neutralize the seasonal variations of hormonal profiles throughout the period, the photoperiod was controlled every 30 days altering long (16 hours of light and 8 hours of darkness) and short days (8 hours of light and 16 hours of darkness). The blood samples were collected twice a week during five weeks. In both breeds, there was no effect of HS (P>0.05) on plasma concentrations of testosterone (1.8±0.2 vs 1.3±0.2ng/ml) and T4 (52.7±2.8 vs 50.0±2.8ng/ml). There was a decline (P<0.01) of plasma T3 concentrations (1.3±0.1 vs 1.0±0.1ng/ml) after HS treatment, but this reduction was only evident in Saanen bucks. In both breeds, the plasma concentrations of T3 and T4 varied (P<0.01) according to the day of blood sample collection. The HS was sufficient to provoke a physiological response with reduction of plasma concentrations of thyroid hormones mainly of T3 in Saanen bucks, but not in Alpine ones. The HS did not affect the plasma testosterone and T4 levels.
buck; heat stress; testosterone; thyroid hormones
MEDICINA VETERINÁRIA
Concentrações plasmáticas de testosterona, triiodotironina (T3) e tiroxina (T4) em bodes submetidos ao estresse calórico
Plasma concentrations of testosterone, triiodothyronine (T3), and thyroxine (T4) in bucks submitted to heat stress
L.A. CoelhoI, * * E-mail: liac@usp.com.br ; A. SasaI; S.D. BicudoII; J.C.C. BalieiroI
IFaculdade de Zootecnia e Engenharia de Alimentos - USP, Av. Duque de Caxias Norte, 225, 13630-000 - Pirassununga, SP
IIFaculdade de Medicina Veterinária e Zootecnia - UNESP - Botucatu, SP
RESUMO
Para verificar o efeito do estresse calórico (EC) nas concentrações plasmáticas de testosterona, triiodotironina (T3) e tiroxina (T4), oito bodes, das raças Saanen (n=4) e Alpina (n=4), foram mantidos em câmara bioclimática, sob condições de termoneutralidade (13,0ºC a 26,7ºC) durante 30 dias e, após um período (60 dias) de descanso, submetidos ao EC (23,7ºC a 34,0ºC) por 30 dias. Para minimizar as variações sazonais nos perfis hormonais devido ao fotoperíodo, durante toda fase experimental, incluindo a de adaptação em condições de termoneutralidade (30 dias), o fotoperíodo foi controlado utilizando-se alternância de dias longos (16h de luz e 8h de escuro) e de dias curtos (8h de luz e 16h de escuro) a cada 30 dias. As amostras de sangue foram coletadas duas vezes por semana durante cinco semanas. No conjunto das raças, o EC não influenciou (P>0,05) as concentrações de testosterona (1,8±0,2 vs 1,3±0,2ng/ml) e nem a de T4 (52,7±2,8 vs 50,0±2,8ng/ml). Houve declínio (P<0,01) das concentrações de T3 nos animais submetidos ao experimento (1,3±0,1 vs 1,0±0,1ng/ml), mas a redução foi observada somente nos bodes Saanen. Em ambas as raças, as concentrações de T3 e T4 variaram (P<0,01) conforme o dia da coleta das amostras de sangue. O EC foi suficiente para produzir uma resposta fisiológica com redução das concentrações plasmáticas de T3 em bodes das raças Saanen, mas não da raça Alpina, assim como não foi capaz de alterar os níveis plasmáticos de testosterona e nem de T4.
Palavras-chave: bode, estresse calórico, hormônios tireoidianos, testosterona
ABSTRACT
To verify the effect of heat stress (HS) on plasma testosterone, triiodothyronine (T3), and thyroxine (T4) concentrations, eight Saanen (n=4) and Alpine Brown (n=4) bucks were kept in climate chamber under thermal neutral conditions (13.0ºC to 26.7ºC) for 30 days. After a resting period (60 days), the same bucks were submitted to heat stress (23.7ºC to 34.0ºC) for another 30 days. To neutralize the seasonal variations of hormonal profiles throughout the period, the photoperiod was controlled every 30 days altering long (16 hours of light and 8 hours of darkness) and short days (8 hours of light and 16 hours of darkness). The blood samples were collected twice a week during five weeks. In both breeds, there was no effect of HS (P>0.05) on plasma concentrations of testosterone (1.8±0.2 vs 1.3±0.2ng/ml) and T4 (52.7±2.8 vs 50.0±2.8ng/ml). There was a decline (P<0.01) of plasma T3 concentrations (1.3±0.1 vs 1.0±0.1ng/ml) after HS treatment, but this reduction was only evident in Saanen bucks. In both breeds, the plasma concentrations of T3 and T4 varied (P<0.01) according to the day of blood sample collection. The HS was sufficient to provoke a physiological response with reduction of plasma concentrations of thyroid hormones mainly of T3 in Saanen bucks, but not in Alpine ones. The HS did not affect the plasma testosterone and T4 levels.
Keywords: buck, heat stress, testosterone, thyroid hormones
INTRODUÇÃO
A elevada temperatura ambiental pode interferir negativamente na temperatura testicular e, conseqüentemente, na qualidade espermática de caprinos, sendo que as características seminais mais afetadas são; a motilidade individual progressiva e o percentual de células morfologicamente anormais (Chemineau, 2004; Coelho et al., 2006). Esse quadro, conhecido como estresse calórico, também provoca algumas alterações nas concentrações plasmáticas de testosterona (Marai et al., 2002) e nos hormônios tireoidianos (Renaudeau et al., 2003; Starling et al., 2005) em mamíferos.
Discrepâncias na literatura, com relação às alterações dos hormônios reprodutivos no macho em função do estresse calórico, têm sido relatadas. Embora existam evidências de que há declínio dos níveis plasmáticos de testosterona em coelhos (Marai et al., 2002), estudo realizado com touros tem demonstrado que o estresse calórico não exerce qualquer efeito deletério sobre as concentrações sangüíneas de testosterona (Kraemer, 2000). Variações nas concentrações de testosterona devido ao estresse calórico em machos caprinos ainda não foram estudadas. Nessa espécie, as pesquisas existentes têm avaliado apenas o efeito do fotoperíodo sobre os hormônios reprodutivos (Delgadillo et al., 2004).
Da mesma forma, os resultados encontrados na literatura sobre alterações do perfil plasmático de triiodotironina (T3) e tiroxina (T4) em diversas espécies domésticas, submetidas ao estresse calórico, têm sido pouco conclusivos. Algumas pesquisas realizadas com animais domésticos (Renaudeau et al., 2003; Starling et al., 2005) submetidos ao estresse calórico têm relatado decréscimo das concentrações plasmáticas de T3 e T4, enquanto outras não conseguiram mostrar qualquer alteração hormonal (Berbegier e Cabelo, 1990; Baccari Jr. et al., 1996; Uribe-Velásquez et al., 1998). Além disso, a maioria dessas pesquisas tem se restringido ao estudo das concentrações plasmáticas de T3 e T4 em fêmeas em lactação (Berbegier e Cabelo, 1990; Baccari Jr., et al., 1996), cujas alterações plasmáticas desses hormônios podem interagir com o metabolismo energético da glândula mamária das fêmeas (Renaudeau et al., 2003), visto que existe uma relação negativa entre produção de leite e concentrações plasmáticas desses hormônios (Vanjonack e Johnson, 1975).
Este trabalho teve como objetivo verificar possíveis variações das concentrações plasmáticas de testosterona, T3 e T4 em machos caprinos das raças Saanen e Alpina, submetidos ao estresse calórico em câmara bioclimática.
MATERIAL E MÉTODOS
O trabalho foi realizado na Câmara Bioclimática do Laboratório de Biometeorologia e Etologia da USP, Campus de Pirassununga-SP. Foram utilizados oito machos, sendo quatro da raça Alpina e quatro da raça Saanen, entre 12 e 18 meses de idade, mantidos em gaiolas metabólicas individuais com 2m2/animal, piso de estrado plástico e acesso ao cocho e bebedouro. As médias dos pesos corporais e escores de condição corporal foram 79,1±1,5kg e 3,2±0,5 e 80,2±1,0 e 3,4±0,2 para animais Alpinos e Saanen, respectivamente. A dieta consistia de feno de coast-cross (Cynodon dactylon), fornecida duas vezes ao dia, sal mineral para caprinos à vontade e um concentrado; 250g/animal/dia, com 22% de proteína bruta, uma vez ao dia, conforme os requisitos nutricionais para caprinos em reprodução (The nutrition..., 1998).
O procedimento experimental teve um período de adaptação de 30 dias, também em condições de termoneutralidade, para os animais se ajustarem às condições físicas e de alimentação nas gaiolas dentro da câmara bioclimática. A seguir, os dados foram obtidos durante 30 dias, em condições de termoneutralidade. Após esse período, promoveu-se um descanso de 60 dias, sem coletas das amostras de sangue, após o qual os dados foram obtidos por mais 30 dias em condições de estresse calórico das 8 às 17horas.
Durante a fase de adaptação e em todo o período experimental, dentro da câmara bioclimática, os valores médios, mínimos e máximos, referentes à temperatura ambiente e à umidade do ar, foram registrados por meio de termoigrógrafos, higrômetros e termômetros de leitura instantânea. As médias mínimas e máximas referentes à temperatura radiante também foram mensuradas por meio de termômetros de globo negro (Tab. 1).
Para minimizar as variações sazonais nas concentrações plasmáticas de testosterona devido ao fotoperíodo (Pérez e Mateos, 1995), estas foram controladas por meio de timer com fornecimento de luz em esquema de alternância de dias longos (16 horas de luz artificial e oito horas de escuro) e de dias curtos (16 horas de escuro e oito horas de luz artificial), a cada 30 dias (Delgadillo et al., 1992). O controle fotoperíodico dentro da câmara bioclimática foi realizado durante os cinco meses do período experimental (período de adaptação, período de coletas dos dados e período sem coleta dos dados).
Após o período de adaptação, amostras de sangue foram obtidas por punção da veia jugular externa, duas vezes por semana, sempre pela manhã, durante cinco semanas. Após centrifugação (1600 G), durante 20 minutos, o plasma sangüíneo foi separado, estocado em frascos plásticos, devidamente identificados, e então acondicionados à temperatura de -20ºC até o momento de serem efetuadas as análises.
As concentrações plasmáticas de testosterona e de T3 e T4 foram analisadas, em duplicatas, pelo método de radioimunoensaio (RIE) em fase sólida. Para tal, foram utilizados conjuntos de reagentes comerciais1 1 COAT-A-COUNT - Diagnostic Products Company, Los Angeles, EUA. , desenvolvidos para avaliação quantitativa dos hormônios, sem qualquer tipo de extração química e processo de purificação, valendo-se do iodo125 (I125) como elemento radioativo traçador. Os procedimentos utilizados foram os especificados pelo fabricante. As contagens de radioatividade foram obtidas pela utilização de contador gama PACKARD, modelo 5000, calibrado automaticamente para I125.
Os dados foram avaliados mediante análise de variância, utilizando-se o procedimento PROC MIXED do SAS (User's..., 1995), com opção REPEATED, seguido pelo teste Tukey para estabelecer a comparação entre médias. Foram considerados os efeitos principais do tratamento (termoneutralidade e estresse calórico) da raça (Alpina e Saanen), do dia da coleta das amostras de sangue e das interações entre os fatores. Os dados referentes ao ambiente térmico, durante o período experimental, foram analisados mediante o procedimento one-way ANOVA, seguido pelo teste Duncan para estabelecer a comparação entre médias.
RESULTADOS E DISCUSSÃO
O perfil plasmático de testosterona não foi afetado (P>0,05) pelo estresse calórico (Tab. 2). Os valores médios das concentrações plasmáticas de testosterona foram 1,83±0,20ng/ml e 1,33±0,20ng/ml para machos submetidos à termoneutralidade e ao estresse calórico, respectivamente. Houve discreta variação (P<0,07) dos níveis plasmáticos em função da raça. Os machos da raça Saanen apresentaram médias mais elevadas, 1,96±0,20ng/ml, que os machos da raça Alpina, 1,20ng/ml. Estes resultados sugerem que o estresse calórico não foi suficiente para alterar a testosterona na circulação periférica de bodes expostos à temperatura média de 34,0±1,3ºC, embora os níveis plasmáticos durante as duas primeiras semanas de modificações nas condições ambientais tenham sido menores (P<0,05) que os observados nos animais submetidos à termoneutralidade (Fig. 1). Estes resultados assemelham-se aos de algumas pesquisas realizadas com bovinos (Minton et al., 1981; Kraemer, 2000), que não foram capazes de mostrar qualquer alteração no perfil de testosterona devido ao estresse calórico, mas diferem de outras que demonstraram redução transitória da concentração sangüínea de testosterona em carneiros (Gomes et al., 1971), touros (Rhynes e Ewing, 1973) e cachaços (Wettemann e Desjardins, 1979). Embora existam evidências de que a resposta testicular ao estresse calórico seja variável conforme a espécie, raça, linhagem e indivíduo (Brito et al., 2004; Setchell, 2006), esse parâmetro não deve ser utilizado como indicativo de uma resposta fisiológica associada ao estresse calórico, visto que machos cujas características quanti-qualitativas do ejaculado foram afetadas negativamente pela elevação da temperatura não apresentaram qualquer alteração hormonal (Minton et al., 1981). Este estudo sugere que essa prerrogativa também é válida para a espécie caprina.
De fato, variações nas concentrações plasmáticas de testosterona nem sempre estão relacionadas com a atividade reprodutiva dos machos (D´Occhio e Brooks, 1983). Segundo Blackshaw et al. (1973), temperaturas elevadas têm efeito deletério na espermatogênese de ratos devido à perda de células germinativas. Entretanto, a apoptose testicular induzida pelo estresse calórico atua por um mecanismo fisiológico independente daquele observado, quando da apoptose testicular induzida por depleção dos níveis de testosterona (Yin et al., 1998).
Adicionalmente, o estresse calórico produzido por diferentes técnicas tem afetado de forma diferenciada as células testiculares (Setchell, 2006). A maioria das técnicas tem mostrado que o estresse calórico afeta principalmente os espermatócitos primários e as espermátides (Vigodner et al., 2003), embora existam evidências de seu efeito sobre as espermatogônias (Zhang et al., 2002) e sobre as células de Sertoli (Zhang et al., 2004). Contudo, não existem evidências de que as células de Leydig e a produção de andrógenos sejam diretamente afetadas (Setchell, 2006).
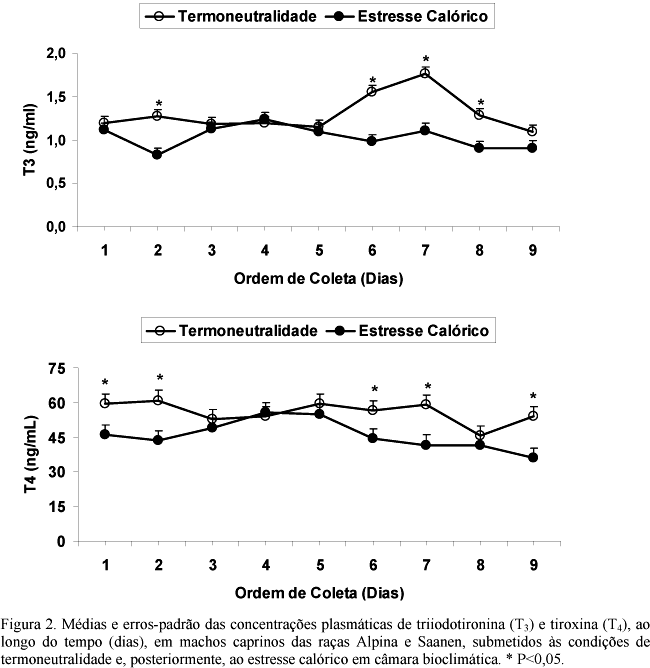
O estresse calórico promoveu significativas alterações na concentração plasmática de T3, mas não na de T4 no conjunto das raças (Tab. 3). Houve declínio (P<0,01) dos níveis plasmáticos de T3 nos machos submetidos ao estresse calórico quando comparados com os machos sob termoneutralidade, entretanto, essa redução foi observada somente nos machos da raça Saanen. Nos machos da raça Alpina, o perfil plasmático de T3 não foi alterado (P>0,05) pelo estresse. Os valores médios obtidos foram 1,29±0,08ng/ml e 0,97±0,08ng/ml na raça Saanen e 1,30±0,08ng/ml e 1,10±0,08 na raça Alpina, para animais em condições de termoneutralidade e estresse calórico, respectivamente. Não houve influência (P>0,05) do estresse calórico e nem da raça nas concentrações plasmáticas de T4 (Tab. 3). As médias oscilaram entre 58,37±3,45ng/ml (Pardo-Alpina) e 50,29±3,45ng/ml (Saanen) para animais mantidos sob termoneutralidade e 49,14±3,45ng/ml (Alpina) e 45,42±3,45ng/ml (Saanen) para animais sob estresse calórico.
O perfil plasmático de ambos os hormônios foi modificado (P<0,01) conforme o dia da coleta das amostras de sangue (Fig. 2). As concentrações plasmáticas de T3 e T4, nos animais estressados, foram reduzidas (P<0,01) em relação aos níveis hormonais dos animais mantidos sob termoneutralidade no início (dia 2 para T3 e dias 1 e 2 para T4) e no final (dias 6, 7 e 8 para T3 e dias 6, 7 e 9 para T4) do período de coleta das amostras de sangue. No início do tratamento, os animais estressados produziram uma resposta fisiológica reduzindo as concentrações médias dos hormônios tireoidianos (Fig. 2), mas parece ter havido adaptação dos animais às condições ambientais durante os terceiro, quarto e quinto dias de tratamento, quando os níveis hormonais foram semelhantes para animais termoneutros e estressados (Fig. 2). A partir daí, os animais voltaram a produzir uma resposta fisiológica à elevação da temperatura ambiental.
Estes resultados assemelham-se aos de outras pesquisas, que têm demonstrado a existência de relação inversa entre as concentrações de hormônios tireoidianos e a temperatura ambiental em cabras (Sano et al., 1985), vacas (Vanjonack e Johnson, 1975; Magdub et al., 1982) e porcas (De Bragança et al., 1998; Renaudeau et al., 2003) lactantes, e em machos e fêmeas da espécie ovina (Starling et al., 2005). Entretanto, diferem dos obtidos por Berbegier e Cabello (1990) com caprinos jovens, cujas mães foram submetidas ao estresse calórico, e por Uribe-Velásquez et al. (1998) com cabras Alpinas adultas, que não observaram qualquer alteração dos perfis plasmáticos desses hormônios decorrente do estresse calórico. A discrepância entre os resultados obtidos e a literatura deve-se principalmente às diferenças dos procedimentos adotados para provocar o estresse calórico (Berbegier e Cabello, 1990) e à freqüência de coleta de amostras de sangue para dosagens hormonais (Uribe-Velásquez et al., 1998).
Os hormônios da tireóide exercem um papel importante no controle da produção de calor em animais homeotérmicos (McNabb, 1995). O decréscimo das concentrações de T3 em resposta ao estresse calórico pode atuar como um mecanismo adaptativo para reduzir a produção de calor (De Bragança et al., 1998). Nesta pesquisa, o estresse calórico foi suficiente para provocar alterações nas concentrações plasmáticas de T3 nos machos da raça Saanen, mas não nos da raça Alpina. Este resultado demonstra que os machos Saanen são menos tolerantes à elevação da temperatura ambiente que os da raça Alpina, concordando com os resultados obtidos em pesquisa anterior realizada com os mesmos animais, nas mesmas condições (Coelho et al., 2006) e com pesquisa realizada com a espécie bovina, que também demonstrou diferenças raciais na tolerância ao estresse calórico (Pegorer et al., 2007).
Neste trabalho, o estresse calórico foi suficiente para produzir uma resposta fisiológica com redução das concentrações plasmáticas de T3 em bodes das raças Saanen, mas não da raça Alpina. Contudo, o estresse calórico não foi capaz de alterar os níveis plasmáticos de testosterona e nem de T4 nos animais de ambas as raças.
Recebido em 22 de fevereiro de 2008
Aceito em 29 de outubro de 2008
Apoio: FAPESP
- BACCARI Jr., F.; GONÇALVES, H.C.; MUNIZ, L.M.R. et al. Milk production, serum concentrations of thyroxine and some physiological responses of Saanen native goats during thermal stress. Rev. Vet. Zootec., v.8, p.9-14, 1996.
- BERBIGIER, P.; CABELLO, G. Effect of exposure to full sunshine on temperature regulation of pregnant dwarf goats of Guadaloupe (French West Indies), and on birthweight, T3 and T4 plasma levels of newborn kids. J. Thermal Biol, v.15, p.109-113, 1990.
- BLACKSHAW, A.W.; HAMILTON, D.J.; MASSEY, P.E. Effects of scrotal heating on testicular enzymes and spermatogenesis in the rat. Aust. J. Biol. Sci., v.26, p.1395-1407, 1973.
- BRITO, L.F.C.; SILVA, A.E.D.F.; BARBOSA, R.T. et al. Testicular thermoregulation in Bos indicus, crossbred and Bos taurus bulls: relationship with scrotal, testicular vascular cone and testicular morphology, and effects on semen quality and sperm production. Theriogenology, v.61, p.511-528, 2004.
- CHEMINEAU, P. Medio ambiente y reproducción animal. World Animal Review, 1993. Disponível em: <http://www.fao.org/DOCREP/V1650T/v1650T04.htm>. Acessado em: 5 jan. 2007.
- COELHO, L.A.; SASA, A.; NADER, C.E. et al. Características do ejaculado de caprinos sob estresse calórico em câmara bioclimática. Arq. Bras. Med. Vet. Zootec, v.58, p.544-549, 2006.
- DE BRAGANÇA, M.M.; MOUNIER, A.M.; PRUNIER, A. Does feed restriction mimic the effects of ambient temperature in lactating sows? J. Anim. Sci, v.76, p.2017-2024, 1998.
- DELGADILLO, J.A.; FITZ-RODRIGUEZ, G.; DUARTE, G. et al. Management of photoperiod to control caprine reproduction in the subtropics. Reprod. Fertil. Develop., v.16, p.471-478, 2004.
- DELGADILLO, J.A.; LEBOEUF, B.; CHEMINEAU, P. Abolition of seasonal variations in semen quality and maintenance of sperm fertilizing ability by photoperiodic cycles in goat bucks. Small Rum. Res., v.9, p.47-59, 1992.
- D´OCCHIO, M.J.; BROOKS, D.E. Seasonal changes in plasma testosterone concentration and mating activity in Border Leicester, Poll Dorset, Romney and Suffolk rams. Aust. J. Exp. Agric. Anim. Husb, v.23, p.248-253, 1983.
- GOMES, W.R.; BUTLER, W.R.; JOHNSON, A.D. Effect of elevated ambient temperature on testis and blood levels and in vitro biosynthesis of testosterone in ram. J. Anim. Sci, v.33, p.804-807, 1971.
- KRAEMER, T. The influence of high ambient temperature on different parameters of semen, biochemical and endocrine parameters in bulls in a climatic chamber and in subtropical climate Berlin: FU Berlin, 2000. Disponível em: <http://www.diss.fu-berlin.de/2000/83/indexe.htm>. Acessado em 5 jan. 2007.
- MAGDUB, A.; JOHNSON, H.D.; BELYEA, R.L. Effect of environmental heat and dietary fiber on thyroid physiology of lactating cows. J. Dairy Sci., v.65, p.2323-2331, 1982.
- MARAI, I.F.M.; HABEEB, A.A.M.; GAD, A.E. Rabbits' productive, reproductive and physiological performance traits as affected by heat stress: a review. Liv. Prod. Sci., v.78, p.71-90, 2002.
- McNABB, A.F.M. Thyroid hormones, their activation, degradation and effects on metabolism. In: CONFERENCE METABOLIC MODIFIERS, 1995, Amsterdam. Proceedings..Amsterdam: Elsevier, 1995. p.1773S-1776S.
- MINTON, J.E.; WETTEMANN, R.P.; MEYERHOFFER, D.C. et al. Serum luteinizing hormone and testosterone in bulls during exposure to elevated ambient temperature. J. Anim. Sci., v.53, p.1551-1558, 1981.
- PEGORER, M.F.; VASCONCELOS, J.L.M.; TRINCA, L.A. et al. Influence of sire and sire breed (Gyr versus Hosltein) on establishment of pregnancy and embryonic loss in lactating Hosltein cows during summer heat stress. Theriogenology, v.67, p.692-697, 2007.
- RENAUDEAU, D.; NOBLET, J.; DOURMAD, J.Y. Effect of ambient temperature on mammary gland metabolism in lactating sows. J. Anim. Sci., v.81, p.217-231, 2003
- PÉREZ, B.; MATEOS, E. Seasonal variations in plasma testosterone levels in Verata and Malagueña bucks. Small Rum. Res., v.15, p.155-162, 1995.
- RHYNES, W.E.; EWING, L.L. Testicular endocrine function in Hereford bulls exposed to high ambient temperature. Endocrinology, v.92, p.509-515, 1973.
- SANO, H.; AMBO, K., TSUDA, T. Blood glucose kinetics in whole body and mammary gland of lactating goats exposed to heat. J. Dairy Sci., v.68, p.2557-2564, 1985.
- SETCHELL, B.P. The effects of heat on testes of mammals. Anim. Reprod., v.3, p.81-91, 2006.
- STARLING, J.M.C.; SILVA, R.G.; NEGRÃO, J.A. et al. Variação estacional dos hormônios tireoideanos e do cortisol em ovinos em ambiente tropical. Rev. Bras. Zootec., v.34, p.2064-2063, 2005.
- THE NUTRITION of goat. AFRC. Wallingford: Cab International, 1998. 118p.
- URIBE-VELÁSQUEZ, L.F.; OBA, E.; BRASIL, L.H.A. Concentrações plasmáticas de cortisol, hormônios tiroídeanos, metabólitos lipídicos e temperatura de cabras Pardo-Alpinas submetidas ao estresse térmico. Rev. Bras. Zootec., v.27, p.1123-1130, 1998.
- USER'S guide: statistics. Cary, NC: SAS Institute, 1995.
- VANJONACK, W.J.; JOHNSON, H.D. Effects of moderate heat and milk yield on plasma thyroxine in cattle. J. Dairy Sci., v.58, p. 507-511, 1975.
- VIGODNER, M.; LEWIN, L.M.; SCHOCHAT, L. et al. Evaluation of damage to the testicular cells of golden hamsters caused by experimental cryptorchidism using flow cytometry and confocal microscopy. Int. J. Androl., v.26, p.84-90, 2003.
- WETTEMANN, R.P.; DESJARDINS, C. Testicular function in boars exposed to elevate ambient temperature. Biol. Reprod, v.20, p.235-241, 1979.
- YIN, Y.; STAHL, B.C.; DeWOLF, W.C. et al. Heat-induced testicular apoptosis occurs independently of hormonal depletion. Apoptosis, v.3, p.281-287, 1998.
- ZHANG, Z.U.; HU, Z.Y.; SONG, X.X. et al. Disrupted expression of intermediate filaments in the testis of the rhesus monkey after experimental cryptorchidism. Int. J. Androl., v.27, p.234-239, 2004.
- ZHANG, R.D.; WEN, X.H.; JONG, L.S. et al. A quantitative (stereological study of the effects of experimental cryptorchidism and subsequent orchidopexy on spermatogenesis in adult rabbit testis. Reproduction, v.124, p.95-105, 2002.
Datas de Publicação
-
Publicação nesta coleção
09 Fev 2009 -
Data do Fascículo
Dez 2008
Histórico
-
Aceito
29 Out 2008 -
Recebido
22 Fev 2008