
Resumo
Novos estudos arqueobotânicos mostram que a região da bacia do alto Madeira é uma área onde foram domesticadas várias plantas durante o Holoceno inicial e médio, confirmando o que já havia sido apontado por dados genéticos há anos. No entanto, há menos acúmulo de dados sobre as relações entre pessoas e plantas para as ocupações humanas no Holoceno tardio. Na calha do alto rio Madeira, ocorrem extensos pacotes de terra preta antropogênica associados a populações ceramistas que viviam na região entre 3.000 e 400 anos atrás. Essas populações deixaram uma riqueza de informações que nos permitem enxergar tais relações através de estudos arqueobotânicos. Este artigo relata novos dados microbotânicos dos sítios Teotônio e Santa Paula para propor novas hipóteses sobre os sistemas de manejo efetuados por essas populações. Além de documentar uma proliferação de cultivos domesticados, foi possível sugerir algumas mudanças no uso de plantas ao longo do tempo. Vimos também como os dados arqueobotânicos podem elucidar questões relacionadas à própria formação dos sítios arqueológicos.
Palavras-chave
Arqueobotânica; Fitólitos; Grãos de amido; Sistemas de manejo; Terra preta antrópica; Sudoeste da Amazônia
Abstract
New archaeobotanical studies show that the region of the Upper Madeira River was where several economic plants were brought under domestication during the Early and Middle Holocene, confirming what had been indicated by plant genetics for several years. Little is known, however, about the more recent history of plant-people relationships, during the ceramic occupations of the Late Holocene. Along the upper Madeira river channel, extensive archaeological sites containing anthropogenic dark earths are associated with occupations by ceramic producers that lived in the area between approximately 3,000 and 400 years ago. These populations left a wealth of information that allows us to evaluate these relationships through archaeobotanical studies. This paper presents new microbotanical data from the Teotônio and Santa Paula archaeological sites to proposes new hypotheses about local plant management systems during this period. Apart from documenting a proliferation in domesticates, it was possible to suggest some changes in which plants were used and how over time. We also saw how archaeobotanical data can shed light upon issues related the formation of these archaeological sites.
Keywords
Archaeobotany; Phytoliths; Starch grains; Management systems; Anthropogenic Dark Earth; Southwest Amazonia
INTRODUÇÃO
O presente estudo tem foco nos dados arqueobotânicos das ocupações ceramistas nos sítios Teotônio e Santa Paula, ambos situados na Cachoeira do Teotônio, na calha principal do rio Madeira, Rondônia, Brasil. O sítio Santa Paula foi construído ca. 1.600 anos calibrados antes do presente (cal. A.P.) (Bespalez et al., 2020Bespalez, E., Zuse, S., Pessoa, C., Venere, P., & Santi, J. (2020). Arqueologia no sítio Santa Paula, alto Madeira, Porto Velho, Roraima, Brasil. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, 15(2), e20190076. doi: 10.1590/2178-2547-BGOELDI-2019-0075
https://doi.org/10.1590/2178-2547-BGOELD...
), enquanto o sítio Teotônio tem indícios de ocupações humanas que recuam até ca. 9.000 cal. A.P. (Watling et al., 2018Watling, J., Shock, M. P., Mongeló, G. Z., Almeida, F. O., Kater, T., De Oliveira, P. E., & Neves, E. G. (2018). Direct archaeological evidence for Southwestern Amazonia as an early plant domestication and food production centre. PLos ONE, 13(7), 1-28. doi: 10.1371/journal.pone.0199868
https://doi.org/10.1371/journal.pone.019...
; Mongeló, 2020Mongeló, G. Z. (2020). Ocupações humanas do Holoceno inicial e médio no sudoeste amazônico. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, 15(2), e20190079. doi: 10.1590/2178-2547-BGOELDI-2019-0079
https://doi.org/10.1590/2178-2547-BGOELD...
), e ocupações ceramistas mais diversas do que as encontradas no Santa Paula (Kater, 2020Kater, T. (2020). A temporalidade das ocupações ceramistas no sítio Teotônio. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, 15(2), e20190078. doi: 10.1590/2178-2547-BGOELDI-2019-0078
https://doi.org/10.1590/2178-2547-BGOELD...
). O nosso objetivo é trazer observações preliminares sobre mudanças e continuidades nas relações entre pessoas e plantas entre aproximadamente 3.000 e 400 cal. A.P. Entre as questões abordadas, estão: i) Podemos sugerir mudanças no uso de plantas (como em quais das plantas foram cultivadas e quais os contextos de consumo e de uso) ao longo da sequência de ocupações ceramistas no Teotônio?; ii) Como o registro arqueobotânico de Santa Paula se relaciona com aquele do Teotônio?; iii) O que os vestígios botânicos podem nos dizer sobre a maneira de ocupar a paisagem, incluindo a própria formação dos sítios?
Antes de nos profundarmos nestas problemáticas, será abordado o contexto arqueológico antes de 3.000 cal. A.P. no alto Rio Madeira e nos próprios sítios Teotônio e Santa Paula.
PLANTAS E OCUPAÇÕES PRÉ-CERÂMICAS NO ALTO MADEIRA: UM RESUMO
A região do alto rio Madeira se destaca dentro do contexto amazônico não apenas por ser altamente rica em sítios arqueológicos, bem como por sua cronologia de ocupação humana recuada, mas, sobretudo, por ter sido um dos maiores centros de domesticação de plantas na América do Sul (Piperno, 2011Piperno, D. R. (2011). The origins of plant cultivation and domestication in the New World tropics: patterns, process, and new developments. Current Anthropology, 52(S4), S453–S470. doi: 10.1086/659998
https://doi.org/10.1086/659998...
; Clement et al., 2016Clement, C. R., Rodrigues, D. P., Alves-Pereira, A., Mühlen, G. S., Cristo-Araújo, M., Moreira, P. A., . . . Reis, V. M. (2016). Crop domestication in the upper Madeira River basin. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, 11(1), 193-205. doi: 10.1590/1981.81222016000100010
https://doi.org/10.1590/1981.81222016000...
; Watling et al., 2018Watling, J., Shock, M. P., Mongeló, G. Z., Almeida, F. O., Kater, T., De Oliveira, P. E., & Neves, E. G. (2018). Direct archaeological evidence for Southwestern Amazonia as an early plant domestication and food production centre. PLos ONE, 13(7), 1-28. doi: 10.1371/journal.pone.0199868
https://doi.org/10.1371/journal.pone.019...
). A mandioca (Manihot esculenta), cujos tubérculos são consumidos hoje em dia por 500 milhões de pessoas nas regiões tropicais do planeta, foi domesticada dentro da mesma macrorregião – ou seja, na área que abrange o sul de Rondônia e as vizinhas terras baixas bolivianas – entre 10 e 8.000 anos atrás (Olsen & Schaal, 1999Olsen, K. M., & Schaal, B. A. (1999). Evidence on the origin of cassava - phylogeography of Manihot esculenta. Proceedings of the National Academy of Sciences of the United States of America, 96(10), 5586-5591. doi: 10.1073/pnas.96.10.5586
https://doi.org/10.1073/pnas.96.10.5586...
; Alves-Pereira et al., 2018Alves-Pereira, A., Clement, C. R., Picanço-Rodrigues, D., Veasey, E. A., Dequigiovanni, G., Ramos, S. L. F., . . . Zucchi, M. I. (2018). Patterns of nuclear and chloroplast genetic diversity and structure of manioc along major Brazilian Amazonian rivers. Annals of Botany, 121(4), 625-639. doi: 10.1093/aob/mcx190
https://doi.org/10.1093/aob/mcx190...
), e em 7.000 cal. A.P. foi incorporada às dietas em regiões muito distantes da América do Sul (Dickau et al., 2007Dickau, R., Ranere, A. J., & Cooke, R. G. (2007). Starch grain evidence for the preceramic dispersals of maize and root crops into tropical dry and humid forests of Panama. Proceedings of the National Academy of Sciences, 104(9), 3651-3656. doi: 10.1073/pnas.0611605104
https://doi.org/10.1073/pnas.0611605104...
; Piperno & Dillehay, 2008Piperno, D. R., & Dillehay, T. D. (2008). Starch grains on human teeth reveal early broad crop diet in northern Peru. Proceedings of the National Academy of Sciences, 105(50), 19622-19627. doi: 10.1073/pnas.0808752105
https://doi.org/10.1073/pnas.0808752105...
). Pelo menos duas variedades de pupunha, a única palmeira a ser inteiramente domesticada (ou seja, com populações que dependem dos seres humanos exclusivamente para sua reprodução), também foram desenvolvidas nessa região e distribuídas pelas terras baixas amazônicas, em diferentes épocas do Holoceno (Clement et al., 2017Clement, C. R., Cristo-Araújo, M., Coppens d’Eeckenbrugge, G., Reis, V. M., Lehnebach, R., & Picanço-Rodrigues, D. (2017). Origin and dispersal of domesticated peach palm. Frontiers in Ecology and Evolution, 5, 1-19. doi: 10.3389/fevo.2017.00148
https://doi.org/10.3389/fevo.2017.00148...
). Estudos genéticos sugerem que o amendoim (Arachis hypogea), a pimenta (Capsicum baccatum), o urucum (Bixa orellana) e a taioba (Xanthosoma spp.) podem ter sidos domesticados no alto rio Madeira (Clement et al., 2016Clement, C. R., Rodrigues, D. P., Alves-Pereira, A., Mühlen, G. S., Cristo-Araújo, M., Moreira, P. A., . . . Reis, V. M. (2016). Crop domestication in the upper Madeira River basin. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, 11(1), 193-205. doi: 10.1590/1981.81222016000100010
https://doi.org/10.1590/1981.81222016000...
). O milho (Zea mays), que chegou no sudoeste amazônico vindo do México em ca. 6.500 anos não calibrados antes do presente (A.P.), sofreu uma parte significativa do seu processo de domesticação após a sua incorporação pelos povos desta região (Kistler et al., 2018Kistler, L., Maezumi, S. Y., Gregorio de Souza, J., Przelomska, N. A., Malaquias Costa, F., Smith, O., Loiselle, H., . . . Allaby, R. G. (2018). Multiproxy evidence highlights a complex evolutionary legacy of maize in South America. Science, 362(6420), 1309-1313. doi: http://doi.org/10.1126/science.aav0207
https://doi.org/10.1126/science.aav0207...
). Nos pantanais da fronteira com a Bolívia, uma espécie local de arroz (Oryza sp.) também foi domesticada ca. 4.000 cal. A.P. (Hilbert et al., 2017Hilbert, L., Neves, E. G., Pugliese, F., Whitney, B. S., Shock, M. P., Veasey, E., . . . Iriarte, J. (2017). Evidence for mid-Holocene rice domestication in the Americas. Nature Ecology & Evolution, 1(11), 1693-1698. doi: 10.1038/s41559-017-0322-4
https://doi.org/10.1038/s41559-017-0322-...
).
Watling et al. (2018)Watling, J., Shock, M. P., Mongeló, G. Z., Almeida, F. O., Kater, T., De Oliveira, P. E., & Neves, E. G. (2018). Direct archaeological evidence for Southwestern Amazonia as an early plant domestication and food production centre. PLos ONE, 13(7), 1-28. doi: 10.1371/journal.pone.0199868
https://doi.org/10.1371/journal.pone.019...
publicaram os resultados de análises arqueobotânicas dos contextos mais antigos do sítio Teotônio. Os contextos tratam de ocupações das fases Girau e Massangana: fases arqueológicas regionais, definidas por Miller et al. (1992)Miller, E. T. [outros não especificados]. (1992). Arqueologia nos empreendimentos hidreléctricos da Electronorte: resultados preliminares. Brasília: Electronorte., e que são evidenciadas na área mais alta do terraço fluvial onde está situado o sítio Teotônio (Mongeló, 2020Mongeló, G. Z. (2020). Ocupações humanas do Holoceno inicial e médio no sudoeste amazônico. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, 15(2), e20190079. doi: 10.1590/2178-2547-BGOELDI-2019-0079
https://doi.org/10.1590/2178-2547-BGOELD...
). A fase Girau, que consiste em artefatos líticos depositados em latossolos (solos não modificados por atividades antrópicas), é aqui encontrada em uma camada diretamente abaixo da fase Massangana, caraterizada pela presença de artefatos líticos em uma matriz de solo com caraterísticas de terra preta antropogênica (TPA), datada entre 6.500 e 5.000 cal. A.P. (Mongeló, 2015Mongeló, G. Z. (2015). O Formativo e os modos de produção: ocupações pré-ceramistas no alto Rio Madeira - RO (Mestrado em Arqueologia). Universidade de São Paulo, São Paulo.; Watling et al., 2018Watling, J., Shock, M. P., Mongeló, G. Z., Almeida, F. O., Kater, T., De Oliveira, P. E., & Neves, E. G. (2018). Direct archaeological evidence for Southwestern Amazonia as an early plant domestication and food production centre. PLos ONE, 13(7), 1-28. doi: 10.1371/journal.pone.0199868
https://doi.org/10.1371/journal.pone.019...
). Através de análises de macro e microvestígios botânicos (respectivamente, carvões, fitólitos e grãos de amido), os autores exploraram a hipótese de que a fase Massangana marca a chegada da ‘agricultura incipiente’ na região (Miller et al., 1992Miller, E. T. [outros não especificados]. (1992). Arqueologia nos empreendimentos hidreléctricos da Electronorte: resultados preliminares. Brasília: Electronorte.) ou, possivelmente, um investimento maior no cultivo da mandioca (Arroyo-Kalin, 2010Arroyo-Kalin, M. (2010). The Amazonian formative: crop domestication and anthropogenic soils. Diversity, 2(4), 473-504. doi: 10.3390/d2040473
https://doi.org/10.3390/d2040473...
).
Na fase Girau, foram encontrados vestígios de tubérculos carbonizados junto com fitólitos dos quais Chandler-Ezell et al. (2006)Chandler-Ezell, K., Pearsall, D. M., & Zeidler, J. A. (2006). Root and tuber phytoliths and starch grains document manioc (Manihot escuelenta), arrowroot (Maranta aruniacea), and llerén (Calathea sp.) at the Real Alto site, Ecuador. Economic Botany, 60(2), 103-120. identificaram nas rizomas comestíveis do ariá (Calathea allouia), mas que possivelmente pertencem a outra espécie do mesmo gênero (Calathea sp.). Tais fitólitos foram encontrados nos mesmos depósitos que sementes de goiaba (Psidium sp.), castanha-do-pará (Bertholettia excelsa), leguminosas (Fabaceae), piquiá (Caryocar sp.) e palmeiras (Arecaceae). Esse conjunto de espécies levou Watling et al. (2018)Watling, J., Shock, M. P., Mongeló, G. Z., Almeida, F. O., Kater, T., De Oliveira, P. E., & Neves, E. G. (2018). Direct archaeological evidence for Southwestern Amazonia as an early plant domestication and food production centre. PLos ONE, 13(7), 1-28. doi: 10.1371/journal.pone.0199868
https://doi.org/10.1371/journal.pone.019...
a sugerirem que, entre ca. 9.000 e 6.500 cal. A.P., as populações pioneiras desta região estavam engajadas na coleta e no possível cultivo de raízes e tubérculos, e que já estavam utilizando fogo como uma ferramenta de manejo da vegetação, ao redor do sítio arqueológico.
Na fase Massangana, foram encontrados fitólitos de mandioca, junto com os microvestígios de dois outros cultivos domesticados – abóbora (Cucurbita sp.) e feijão (Phaseolus sp.). Tanto a abóbora quanto o feijão são considerados plantas exóticas, devido às suas origens fora desta região. Watling et al. (2018)Watling, J., Shock, M. P., Mongeló, G. Z., Almeida, F. O., Kater, T., De Oliveira, P. E., & Neves, E. G. (2018). Direct archaeological evidence for Southwestern Amazonia as an early plant domestication and food production centre. PLos ONE, 13(7), 1-28. doi: 10.1371/journal.pone.0199868
https://doi.org/10.1371/journal.pone.019...
sugerem que a própria formação de TPA, durante a fase Massangana, criou solos mais favoráveis que possibilitaram a adoção e experimentação com essas espécies, que têm maior demanda por nutrientes, nos jardins das casas (sensuOliver, 2008Oliver, J. R. (2008). The archaeology of agriculture in ancient Amazonia. In H. Silverman & W. H. Isbell (Orgs.), Handbook of South American Archaeology (pp. 185-216). New York: Springer.), enquanto a coleta de frutas como castanha-do-pará e bacaba (Oenocarpus sp.) continuava sendo importante. Uma discrepância entre as quantidades de fitólitos versus sementes de palmeiras no registro arqueobotânico levou à hipótese de que os fitólitos vieram do material construtivo de casas, ao invés das frutas, corroborando os indícios de que o contexto em questão era uma área doméstica.
No Holoceno médio, portanto, antes do início das ocupações ceramistas, já havia uma população regional de ‘caçadores-coletores-jardineiros’ (sensuKillion, 2013Killion, T. W. (2013). Nonagricultural cultivation and social complexity: the Olmec, their ancestors, and Mexico’s Southern Gulf Lowlands. Current Anthropology, 54(5), 569-606. doi: 10.1086/673140
https://doi.org/10.1086/673140...
) no alto rio Madeira, investida no cultivo de plantas domesticadas.
AS OCUPAÇÕES CERAMISTAS DO ALTO MADEIRA: NOVAS PERGUNTAS
As evidências mais antigas de cerâmicas no alto rio Madeira são de ca. 3.000 cal. A.P., quando foi iniciada uma rica história de ocupações, conhecidas através da presença de várias tradições ceramistas regionais e locais que duraram até o período colonial, ca. 400 cal. A.P. (Zuse, 2014Zuse, S. (2014). Variabilidade cerâmica e diversidade cultural no alto rio Madeira, Rondônia (Doutorado em Arqueologia). Universidade de São Paulo, São Paulo., 2016Zuse, S. (2016). Variabilidade cerâmica e diversidade cultural no alto rio Madeira. In Barreto, C., Lima, H. P., & Jaimes Betancourt, C. (Orgs.), Cerâmicas arqueológicas da Amazônia: rumo a uma nova síntese (pp. 385-401). Belém: IPHAN., 2020Zuse, S. (2020). Tecnologias cerâmicas no alto rio Madeira: síntese, cronologia e perspectivas. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, 15(2), e20190082. doi: 10.1590/2178-2547-BGOELDI-2019-0082
https://doi.org/10.1590/2178-2547-BGOELD...
). A chegada de pessoas produzindo cerâmicas da tradição Pocó-Açutuba entre ca. 3.000 e 2.500 cal. A.P. marca, tanto no rio Madeira quanto na bacia amazônica como um todo, o começo do período Formativo (Neves, 2006Neves, E. G. (2006). Arqueologia da Amazônia. Rio de Janeiro: Jorge Zahar.; Arroyo-Kalin, 2010Arroyo-Kalin, M. (2010). The Amazonian formative: crop domestication and anthropogenic soils. Diversity, 2(4), 473-504. doi: 10.3390/d2040473
https://doi.org/10.3390/d2040473...
). Apesar dos inúmeros problemas em aplicar o termo ‘Período Formativo’ na Amazônia (Neves, 2007Neves, E. G. (2007). El Formativo que nunca terminó: la larga historia de estabilidad en las ocupaciones humanas de la Amazonía Central. Boletín de Arqueología PUCP, 11, 117-142., 2013Neves, E. G. (2013). Was agriculture a key productive activity in pre-Colonial Amazonia? The stable productive basis for social equality in the Central Amazon. In E. S. Brondízio & E. F. Moran (Orgs.), Human-Environment Interactions: current and future decisions (pp. 371-388). Dordrecht: Springer.), os vestígios arqueológicos deixam claro que a partir desse momento ficaram mais visíveis os sinais de criação de paisagens pelas populações indígenas (Neves et al., 2014Neves, E. G. (2014). La incipiencia permanente: la Amazonia bajo el insistente destino de la incompletitud. In M. Campagno (Org.), Pierre Clastres y Las Sociedades Antiguas (1. ed., pp. 65-80). Buenos Aires: Miño y Dávila.). Tais sinais resultam do maior investimento no esforço do cultivo de plantas domesticadas, seja com a mandioca ou o milho como alimentos de base (Lathrap, 1970Lathrap, D. W. (1970). The Upper Amazon. London: Thames and Hudson.; Roosevelt, 1980Roosevelt, A. C. (1980). Parmana: prehistoric maize and manioc subsistence along the Amazon and Orinoco. New York: Academic Press.), ou dentro de sistemas agroflorestais de policultura (Denevan, 2006Denevan, W. M. (2006). Pre-European forest cultivation in Amazonia. In W. Balée & C. L. Erickson (Orgs.), Time and complexity in historical ecology (pp. 153-164). New York: Columbia University Press.; Maezumi et al., 2018Maezumi, S. Y., Alves, D., Robinson, M., Gregorio de Souza, J., Levis, C., Barnett, R. L., . . . Iriarte, J. (2018). The legacy of 4,500 years of polyculture agroforestry in the eastern Amazon. Nature Plants, 4(8), 540-547. doi: 10.1038/s41477-018-0205-y
https://doi.org/10.1038/s41477-018-0205-...
), que dependeria da abundância dos recursos naturais (Neves, 2014Neves, E. G., Guapindaia, V. L. C., Lima, H. P., Costa, B. L. S., & Gomes, J. (2014). A tradição Pocó-Açutuba e os primeiros sinais visíveis de modificações de paisagens na calha do Amazonas. In S. Rostain (Org.), Antes de Orellana: Actas del 3er Encuentro Internacional de Arqueología Amazônica (pp. 137-158). Quito: IFEA.) e da fartura criada por sistemas de manejo de recursos e paisagens (Clement et al., 2015Clement, C. R., Denevan, W. M., Heckenberger, M. J., Junqueira, A. B., Neves, E. G., Texeira, W. G., & Woods, W. I. (2015). The domestication of Amazonia before European conquest. Proceedings of the Royal Society B, 282(1812), 1-9. doi: 10.1098/rspb.2015.0813
https://doi.org/10.1098/rspb.2015.0813...
; Levis et al., 2017Levis, C., Costa, F. R. C., Bongers, F., Peña-Claros, M., Clement, C. R., Junqueira, A. B., . . . Ter Steege, H. (2017). Persistent effects of pre-Columbian plant domestication on Amazonian forest composition. Science, 355(6328), 925-931. doi: 10.1126/science.aal0157
https://doi.org/10.1126/science.aal0157...
).
A transição entre os períodos pré-ceramista e ceramista no alto Madeira ainda está para ser entendida. Vimos, entretanto, que já existiriam TPA e um forte sistema de produção e manejo de recursos vegetais bem antes da chegada dos produtores de cerâmica Pocó-Açutuba. Por outro lado, sabemos que estas populações estão apenas entre as primeiras dentro de uma longa história de ocupações e interações multiétnicas na região (Almeida & Kater, 2017Almeida, F. O., & Kater, T. (2017). As cachoeiras como bolsões de histórias dos grupos indígenas das terras baixas sul-americanas. Revista Brasileira de História, 37(75), 39-67. doi: 10.1590/1806-93472017v37n75-02a
https://doi.org/10.1590/1806-93472017v37...
), as quais podem ter introduzido diversas mudanças nos sistemas de manejo ao longo do tempo.
Neste trabalho, dados do sítio Santa Paula oferecem uma perspectiva sincrônica desses sistemas ao redor de 1.500 cal. A.P. O sítio Teotônio, visto como um microcosmo de ocupações regionais (Almeida & Kater, 2017Almeida, F. O., & Kater, T. (2017). As cachoeiras como bolsões de histórias dos grupos indígenas das terras baixas sul-americanas. Revista Brasileira de História, 37(75), 39-67. doi: 10.1590/1806-93472017v37n75-02a
https://doi.org/10.1590/1806-93472017v37...
), oferece uma oportunidade única de enxergar a diversidade de sistemas de manejo ao longo de uma cronologia que dura quase todo o Holoceno. Porém, um sítio tão complexo como este apresenta inúmeros desafios que derivam da própria reocupação do espaço ao longo do tempo. Por isso, exploramos, como mais uma vertente no presente trabalho, a arqueobotânica como ferramenta para entender a formação do contexto arqueológico.
MATERIAIS E MÉTODOS
CONTEXTOS ANALISADOS NO PRESENTE ESTUDO
No sítio Teotônio, as TPA associadas às ocupações ceramistas são encontradas em uma área de mais de 50 hectares. As amostras coletadas para análises microbotânicas, sumarizadas no Quadro 1, vieram das unidades de escavação N10041 E9955 (cerâmicas) e N10049 E9956 (solos e líticos), ambas localizadas em uma área do sítio que evidenciou TPA com até 4 metros de profundidade e onde cerâmicas Pocó-Açutuba (P-A), Jamari (JM) e Jatuarana (JT) foram encontradas em sobreposição (Figura 1). Com base na estratigrafia e na análise cerâmica da unidade N10049 E9956 (Kater, 2018Kater, T. (2018). O sítio Teotônio e as reminiscências de uma longa história indígena no alto Rio Madeira (Mestrado em Arqueologia). Universidade Federal de Sergipe, Sergipe, Aracajú.; Kater et al., 2020Kater, T., Almeida, F. Almeida, F. O, Mongeló, G. Z.; Watling, J.; Neves, E. G. (2020). Variabilidade estratigráfica e espacial dos contextos cerâmicos no sítio Teotônio. Revista de Arqueologia, 33(1), 198-220.), foi hipotetizado que a mesma representa um aterro cujos dois metros de base – onde se identificou cerâmicas P-A misturadas com fragmentos de outras tradições que cronologicamente são posteriores e/ou contemporâneas – representam uma, ou mais de uma, camada construtiva (Figura 1) (Kater et al., 2020Kater, T. (2020). A temporalidade das ocupações ceramistas no sítio Teotônio. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, 15(2), e20190078. doi: 10.1590/2178-2547-BGOELDI-2019-0078
https://doi.org/10.1590/2178-2547-BGOELD...
). Os materiais JM e JT dessa unidade, que estão sobrepostos às camadas inferiores, apresentam a TPA mais escura no perfil (10YR 2/1) e representariam, conforme essas análises, contextos in situ.
Tabela mostrando dados sobre os contextos analisados neste estudo. Legendas: * = amostras para fitólitos foram retiradas dos perfis das escavações em intervalos de 10 em 10 cm.
Perfis estratigráficos do Teotônio e Santa Paula cujos solos foram analisados para fitólitos.
O sítio Santa Paula (Zuse, 2014Zuse, S. (2014). Variabilidade cerâmica e diversidade cultural no alto rio Madeira, Rondônia (Doutorado em Arqueologia). Universidade de São Paulo, São Paulo.; Bespalez et al., 2020Bespalez, E., Zuse, S., Pessoa, C., Venere, P., & Santi, J. (2020). Arqueologia no sítio Santa Paula, alto Madeira, Porto Velho, Roraima, Brasil. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, 15(2), e20190076. doi: 10.1590/2178-2547-BGOELDI-2019-0075
https://doi.org/10.1590/2178-2547-BGOELD...
) consiste em um arranjo de montículos de TPA, em formato semicircular, cujas estratigrafia e datações evidenciam um só episódio de construção, entre 1.600 e 1.500 cal. A.P. Cerâmicas da tradição P-A, misturadas a fragmentos com características tecnológicas distintas e possivelmente contemporâneas ou mais antigas, foram encontradas na base dos montículos e são anteriores à sua construção, que está associada a cerâmicas da tradição Borda Incisa/Barrancoide (BC). Cerâmicas JT foram encontradas nos níveis mais superficiais dos montículos, misturadas com cerâmicas BC, deixando em aberto se os portadores dessas cerâmicas substituíram ou não as populações anteriores, como parece ser o caso do Teotônio.
EXTRAÇÃO E IDENTIFICAÇÃO DE FITÓLITOS
Fitólitos foram extraídos dos solos utilizando o método de Lombardo et al. (2016)Lombardo, U., Ruiz-Pérez, J., & Madella, M. (2016). Sonication improves the efficiency, efficacy and safety of phytolith extraction. Review of Palaeobotany and Palynology, 235, 1-5. doi: 10.1016/j.revpalbo.2016.09.008
https://doi.org/10.1016/j.revpalbo.2016....
: 15 g de solo foram colocadas em tubos Falcon de 50 ml. Primeiro, foi adicionada uma solução de hexametafosfato de sódio (5%), e a amostra foi deixada em um banho de ultrassom a 60 °C por 15 minutos para dispersar as argilas. Uma parte da argila foi, então, removida através de duas rodadas de centrifugação a 1.500 rpm por 3 minutos, e a amostra foi lavada com água duas vezes. Os carbonatos foram removidos pela adição de 20 ml de HCl (10%), sendo a amostra lavada duas vezes com água e centrifugada a 2.500 rpm por 2 minutos. A amostra foi, então, peneirada para separar as frações de silte (< 53 µm) e areia (53-250 µm), criando duas subamostras. Para retirar o material orgânico, foram adicionados 20 ml de H2O2 (30%) a cada subamostra, antes de colocar no banho de ultrassom, aquecido a 60 °C. Ciclos de 30 minutos com ultrassom ‘ligado’ e 30 minutos de ultrassom ‘desligado’ foram realizados até não haver mais reação (para as terras pretas, esse processo levou em média 7 horas). A amostra, então, foi lavada três vezes com água com centrifugação de 2.500 rpm por 2 minutos. Para remover o resto da argila, foram adicionados 45 ml da solução de hexametafosfato de sódio, e a amostra foi novamente colocada no banho de ultrassom a 60 °C por 15 minutos, centrifugada a 1.500 rpm por 3 minutos, sendo descartada a solução com argila. Esse processo foi repetido até o sobrenadante ficar claro. Para separar os fitólitos do solo, 20 ml de cloreto de zinco (com densidade específica de 2,3 g/cm3) foram adicionados a cada amostra, que foi misturada com misturador vortex e centrifugada a 3.000 rpm por 5 minutos. Uma pipeta descartável foi usada para transferir 3 ml da superfície do líquido para um novo tubo de 15 ml, e água foi adicionada ao tubo para completar. A amostra foi centrifugada a 3.000 rpm por 10 minutos, antes de ser lavada duas vezes com água (2.500 rpm por 2 minutos). Para secar o resíduo, 4 ml de acetona foram adicionados à amostra, que foi centrifugada a 2.500 rpm por 2 minutos, antes da acetona ser descartada. Depois de secar durante a noite, o resíduo foi colocado em uma lâmina de microscopia utilizando Permount.
A observação dos microvestígios foi realizada com magnificação de 200 e 400 vezes no microscópio Zeiss Lab A1. Foram contados 200 fitólitos com significância taxonômica e a porcentagem de cada tipo foi calculada para processamento no programa C2 (Juggins, 2010Juggins, S. (2010). C2 Software (Versão 1.6) [software de computador]. Austrália. Recuperado de https://www.staff.ncl.ac.uk/stephen.juggins/software/C2Home.htm
https://www.staff.ncl.ac.uk/stephen.jugg...
). A identificação de fitólitos foi feita através de comparações com a literatura e a coleção de referência de plantas modernas no Instituto de Geociências da Universidade de São Paulo. Para a descrição, optamos pelo uso do International Code for Phytolith Nomenclature (Madella et al., 2005Madella, M., Alexandre, A., & Ball, T., 2005. International Code for Phytolith Nomenclature 1.0. Annals of Botany, 96(2), 253-260. doi: https://doi.org/10.1093/aob/mci172
https://doi.org/10.1093/aob/mci172...
), que, apesar de manter a nomenclatura das morfologias dos fitólitos em inglês, evita incertezas decorrentes da tradução (e.g. Cascon, 2010Cascon, L. M. (2010). Alimentação na floresta tropical: um estudo de caso no sítio Hatahara, Amazônia Central, com base em microvestígios botânicos (Mestrado em Arqueologia). Universidade Federal do Rio de Janeiro, Rio de Janeiro.).
A diferenciação de fitólitos produzidos por palmeiras (Arecaceae) seguiu o trabalho de Morcote-Rios et al. (2016)Morcote-Ríos, G., Bernal, R., & Raz, L. (2016). Phytoliths as a tool for archaeobotanical, palaeobotanical and palaeoecological studies in Amazonian palms. Botanical Journal of the Linnean Society, 182(2), 348-360. doi: https://doi.org/10.1111/boj.12438
https://doi.org/10.1111/boj.12438...
e considerou as espécies nativas da região. Neste esquema, os globular echinates symmetric são atribuídos à subtribo Mauritinae e ao gênero Euterpe sp.; os globular echinates elongate são atribuídos a Euterpeae e/ou a Attaleinae; os globular echinate with short dense projections são exclusivamente de Euterpeae; e os cones são dos gêneros Bactris e/ou Astrocaryum. Na publicação de Watling et al. (2018)Watling, J., Shock, M. P., Mongeló, G. Z., Almeida, F. O., Kater, T., De Oliveira, P. E., & Neves, E. G. (2018). Direct archaeological evidence for Southwestern Amazonia as an early plant domestication and food production centre. PLos ONE, 13(7), 1-28. doi: 10.1371/journal.pone.0199868
https://doi.org/10.1371/journal.pone.019...
, os autores também separaram fitólitos de Attaleinae baseando-se no morfotipo globular echinate assymetric, de Morcote-Rios et al. (2016), porém um estudo mais profundo do material moderno das palmeiras regionais não mais apoia esta correspondência, já que esse morfotipo é encontrado em outros membros da família (Watling et al., 2020Watling, J., Castro, M. T., Simon, M. F., Rodrigues, F. O., Brilhante de Medeiros, M., De Oliveira, P. E, Neves, E. G. (2020). Phytoliths from native plants and surface soils in the Upper Madeira basin, SW Amazonia, and their potential for paleoecological reconstruction. Quaternary International. doi: https://doi.org/10.1016/j.quaint.2020.03.045.
https://doi.org/10.1016/j.quaint.2020.03...
).
Além da quantificação taxonômica dos fitólitos, cálculos de concentração também foram feitos para uma amostra de todas as principais camadas da unidade N10049 E9956, do sítio Teotônio. O método seguiu Katz et al. (2010)Katz, O., Cabanes, D., Weiner, S., Maeir, A. M., Boaretto, E., & Shahack-Gross, R. (2010). Rapid phytolith extraction for analysis of phytolith concentrations and assemblages during an excavation: an application at Tell es-Safi/Gath, Israel. Journal of Archaeological Science, 37(7), 1557-1563. doi: 10.1016/j.jas.2010.01.016
https://doi.org/10.1016/j.jas.2010.01.01...
: foram homogeneizadas aproximadamente 20-30 mg da fração de solo 0-53 µm, as quais foram transferidas para um tubo Eppendorf de 1 ml. Em seguida, foram adicionados 50 µl de HCl (6N), seguido por 50 µl de SPT (com densidade específica de 2,4 g/cm3), sendo adicionados e misturados em um misturador vortex, para posterior banho de ultrassom por 10 minutos. Depois de ser centrifugado por 5 minutos a 5.000 rpm, o sobrenadante foi transferido para outro tubo Eppendorf, misturado novamente no vortex, e 50 µl foram transferidos para uma lâmina de microscopia. Com um microscópio, o número de fitólitos em 15 campos aleatórios foi somado, e a concentração foi calculada usando-se os seguintes passos: o número de fitólitos na lâmina (nL) = número de fitólitos contados / área da lâmina contada (mm2) x área total da lâmina. Assim, número de fitólitos em 1 g de amostra (a concentração) = [(nL x 10) / peso original da amostra (mg)] x 1.000.
EXTRAÇÃO E IDENTIFICAÇÃO DE GRÃOS DE AMIDO
Os artefatos cerâmicos e líticos analisados neste estudo encontravam-se em diferentes estágios de curadoria, o que resultou na aplicação de diferentes abordagens. Os métodos descritos a seguir estão em acordo com os publicados em The International Code for Starch Nomenclature (2011)The International Code for Starch Nomenclature (ICSN). (2011). Recuperado de http://fossilfarm.org/ICSN/Code.html
http://fossilfarm.org/ICSN/Code.html...
, Torrence e Barton (2006)Torrence, R., & Barton, H. (2006). Ancient Starch Research. Walnut Creek, California: Left Coast Press Inc. e Watling et al. (2018)Watling, J., Shock, M. P., Mongeló, G. Z., Almeida, F. O., Kater, T., De Oliveira, P. E., & Neves, E. G. (2018). Direct archaeological evidence for Southwestern Amazonia as an early plant domestication and food production centre. PLos ONE, 13(7), 1-28. doi: 10.1371/journal.pone.0199868
https://doi.org/10.1371/journal.pone.019...
.
Todos os equipamentos e vidrarias utilizados na extração de grãos de amido foram previamente esterilizados em uma autoclave. A extração aconteceu em um laboratório que tinha sido completamente higienizado no dia anterior.
As cerâmicas do Teotônio já tinham sido lavadas e analisadas antes da extração das amostras. Para remover a potencial contaminação, foram primeiramente lavadas com uma escova de dente, em água destilada. Uma vez secos, os resíduos grudados dentro das imperfeições da pasta, no interior das peças, foram removidos com o uso de agulhas. Esses resíduos foram montados em lâminas de vidro, em uma solução de 50:50 água destilada e glicerina, para análise no microscópio.
As seis lascas de quartzo dos contextos ceramistas do Teotônio não tinham sido lavadas após a sua coleta em campo, o que permitiu uma análise separada do sedimento aderente aos artefatos e do resíduo mais provavelmente ligado ao uso do artefato. O sedimento foi removido dos artefatos com uma escova de dente e água destilada, e o resíduo foi concentrado em um tubo de ensaio através de centrifugação a 2.000 rpm por 5 minutos. Uma gota do resíduo com água foi transferida para uma lâmina com uma gota de glicerina, misturada e selada com uma lamínula, para a análise microscópica. A segunda extração dos artefatos foi feita colocando-os dentro de um béquer de vidro pequeno, cobrindo-os com água destilada e colocando-os no banho de ultrassom por 5 minutos. O resíduo, então, foi concentrado e montado como antes.
As cerâmicas do sítio Santa Paula foram isoladas em campo com a intenção de fazer análises de grãos de amidos. Nesse caso, extraímos amido do sedimento aderido nos artefatos, da mesma forma do que nos líticos, e a extração mais intensiva foi feita através do uso de agulhas nas imperfeições da pasta. Como os artefatos 2314.2, 2006.4 e 2113.6 (Tabela 1) apresentaram muito sedimento, os grãos de amido foram deles separados com cloreto de césio (de densidade específica de 1,8 g/cm3) e lavados com água destilada, antes de serem montados com glicerina.
Detalhamento dos grãos de amido e fitólitos recuperados nos artefatos analisados dos sítios Teotônio e Santa Paula. Legendas: na coluna “Artefato”, Cer = cerâmica; Lit= lítico; Sed = sedimento; quantidades em negrito são os vestígios encontrados na extração mais intensiva do artefato [i.e. no banho de ultrassom (se for lítico) ou extração de agulha (se for cerâmica)]; quantidades que não estão em negrito representam os vestígios encontrados na extração menos intensiva do artefato (i.e. na lavagem com água destilada); quantidades entre parênteses são identificações taxonômicas tentativas. Na coluna “Danos presentes” (observados nos grãos de amido): AH = abertura do hilo; PH = projeções no hilo; F = fissuras; G = gelatinização; para os fitólitos de Arecaceae, x indica a presença de globular echinates e * indica a presença de cones de Bactris/Astrocaryum.
A identificação dos grãos de amido foi feita através de comparações com a literatura estabelecida e com uma coleção de referência de plantas modernas. No microscópio, foram observados tanto sob luz transmitida quanto polarizada, com magnificações de 400 e 630 vezes. Vários grãos de amido apresentaram caraterísticas de danificação adquiridas no processamento dos alimentos. Seguindo a literatura (Babot, 2003Babot, M. P. (2003). Starch grain damage as an indicator of food processing. In D. M. Hart & L. A. Wallis (Orgs.), Phytolith and starch research in the Australian-Pacific-Asian regions: the state of the art (pp. 69-81). Canberra: Pandanus Books.; The International Code for Starch Nomenclature, 2011)The International Code for Starch Nomenclature (ICSN). (2011). Recuperado de http://fossilfarm.org/ICSN/Code.html
http://fossilfarm.org/ICSN/Code.html...
, os seguintes danos foram definidos: aberturas do hilo (AH: a abertura do hilo causada pela perda de água no grão, através de danificação enzimática ou por aquecimento); projeções do hilo (PH: projeções escuras na área do hilum causadas por serem expostas ao calor); fissuras (F: fissuras no hilo ou na margem do grão pelo dano físico, por exemplo por moagem); gelatinização (G: a perda da estrutura do grão do amido através de aquecimento em água).
RESULTADOS
RESÍDUOS DOS ARTEFATOS
Os resultados dos artefatos arqueológicos estão apresentados na Tabela 1.
TEOTÔNIO
A recuperação de grãos de amido nas cerâmicas do Teotônio foi muito alta, alcançando um máximo de 33 grãos por artefato em fragmentos da tradição P-A. Vinte morfotipos diferentes foram observados, porém somente oito puderam ser taxonomicamente identificados.
A espécie mais representada nas cerâmicas foi o milho, sendo encontrada em 12 dos 17 fragmentos cerâmicos analisados das tradições P-A e JM. Grãos de amido produzidos por milho de endospermas mole (farinhosa) e duro (e.g. pipoca) foram encontrados nos mesmos resíduos (Figuras 2A e 2B), implicando a ideia de que diferentes variedades estavam sendo usadas nesse período. Vários dos grãos tinham sofrido danos consistentes com moagem e exposição ao calor (C, HP e G, na Tabela 1), o que indica a produção e o processamento de farinha de milho.
Grãos de amido de abóbora (Cucurbita sp., Figura 2C) também foram recuperados das cerâmicas P-A e JM, junto com um fitólito do gênero (Figura 2D). A espécie de abóbora é provavelmente Cucurbita moschata (Figura 2C) (D. R. Piperno, comunicação pessoal, junho, 2016).
Prancha de alguns microvestígios encontrados no presente estudo: A) grão de amido de milho (endosperma mole); B) grão de amido de milho (endosperma duro); C) grão de amido de abóbora (Cucurbita cf. moschata); D) fitólito de abóbora (Cucurbita sp.); E) aglomeração de grãos de amido de mandioca; F) grão de amido de batata doce; G) grão de amido de uma leguminosa (Fabaceae); H) grão de amido de cará; I) grão de amido de mandioca com fissuras e cruz de extinção fraca, indicando dano físico; J) fitólito de mandioca; K) fitólito UID 1 (não identificado, possivelmente Marantaceae); L) fitólito UID 2 (não identificado, possivelmente Marantaceae); M) fitólito de araruta (Maranta arundinacea); N) fitólito de Calathea sp. (cf. ariá, C. allouia); O) fitólito da espiga de milho. Prancha: Jennifer Watling (2019)Watling, J., Castro, M. T., Simon, M. F., Rodrigues, F. O., Brilhante de Medeiros, M., De Oliveira, P. E, Neves, E. G. (2020). Phytoliths from native plants and surface soils in the Upper Madeira basin, SW Amazonia, and their potential for paleoecological reconstruction. Quaternary International. doi: https://doi.org/10.1016/j.quaint.2020.03.045.
https://doi.org/10.1016/j.quaint.2020.03... .
Três grãos de amido possivelmente pertencentes à mandioca foram encontrados em uma cerâmica JT, os quais, no entanto, foram ocasionalmente encontrados em outras espécies, fazendo com que a identificação seja preliminar. Nas cerâmicas JT, os grãos de amido são menos frequentes do que nas cerâmicas P-A e JM, estando ausentes na maioria dos fragmentos analisados.
Por outro lado, dois dos cinco artefatos líticos analisados (PNs 1803-2 e 3) do contexto JT apresentaram vários grãos de amido diagnósticos, entre eles aglomerações de grãos de amido de mandioca (Figura 2E), batata doce (Ipomoea batatas) (Figura 2F), leguminosas (Fabaceae) (Figura 2G) e, possivelmente, cará (Dioscorea sp.) (Figura 2H). Alguns grãos de amido de milho foram encontrados quase que exclusivamente nas amostras de lavagem, sugerindo que eles vieram do sedimento, ao invés do artefato. Alguns grãos de amido de mandioca exibiram fissuras e cruzes de extinção apagadas (Figura 2I), evidências associadas a prováveis danos físicos sofridos por esses grãos. Uma hipótese é que decorrem do provável uso de raladores para o processamento da mandioca (ver Discussão). A presença de outros danos nos grãos de amido (AH, AP, G) também sugere que alguns deles foram aquecidos antes de serem ralados. Apenas um dos três líticos do contexto JM continha grãos de amido, estes sendo de milho e de duas espécies não identificadas.
Os fitólitos recuperados dos artefatos foram registrados em dados de presença. Todos eles também estiveram presentes nos conjuntos de fitólitos encontrados nos solos dos mesmos contextos (morfotipos arbóreos, Arecaceae, Poaceae e Asteraceae) e não forneceram dados de interesse do ponto de vista alimentício. As exceções foram os fitólitos de abóbora (1806-226) e cf. ariá (1803-1), encontrados em dois dos fragmentos cerâmicos.
SANTA PAULA
As cerâmicas de Santa Paula, todas pertencentes à tradição Barrancoide/Borda Incisa, continham poucos grãos de amido, se comparadas com as do Teotônio. Quase todos os grãos foram identificados como sendo de milho (identificações definitivas e tentativas), e os não identificados incluíram alguns dos mesmos morfotipos encontrados nos líticos do Teotônio (tipos 8 e 13). Fitólitos de mandioca (Figura 2J) foram encontrados na amostra de raspagem em dois dos artefatos (2006.4 e 2314.1), o primeiro contendo dois deste morfotipo.
O fragmento de assador (PN 2113.6), ao contrário do esperado, não foi uma boa fonte de amidos, contendo um grão de amido de milho e outro de cará no sedimento aderente ao artefato, possivelmente não do próprio artefato. Grãos de amido gelatinizados e com hilum openings foram comuns, sendo que o próprio processamento dos alimentos poderia ter levado à destruição de muitos restos. Os fitólitos dos mesmos artefatos, como no Teotônio, vieram da vegetação local.
FITÓLITOS DOS SOLOS
Teotônio
Os resultados da análise estratigráfica da unidade N10049 E9956 do Teotônio estão apresentados nas Figuras 3 e 4, e os dados de concentração de fitólitos constam do Quadro 2. Recomenda-se conferir o Quadro 1 e a Figura 1 durante as discussões das diferentes camadas arqueológicas.
Porcentagens de fitólitos da fração silt (0-53 µm) encontradas na unidade N10049 E9956 de Teotônio.
Contagens absolutas de fitólitos da fração arenosa (53-250 µm) encontrados na unidade N10049 E9956 de Teotônio.
Dados de concentração de fitólitos em amostras das diferentes camadas da unidade N10049 E9956 do Teotônio, comparado com amostras de um perfil de controle localizado 5 km fora do sítio.
Os dados mostram que todas as camadas de ocupação nesta área do sítio, de até 3,8 m de profundidade, têm concentrações de fitólitos parecidas, e que tais concentrações são muito mais elevadas do que nos solos naturais amostrados a 5 km do sítio. A concentração elevada de fitólitos na camada VI E poderia refletir uma deposição mais intensiva de matéria orgânica durante a fase JT. Outra possibilidade é que, por ser a camada mais superficial, os fitólitos poderiam ser oriundos de percolações resultantes da decomposição de plantas mais jovens. Até agora, nenhum estudo foi realizado sobre a percolação de fitólitos em terras pretas, mas vários trabalhos (a exemplo de Alexandre et al., 1999Alexandre, A., Meunier, J.-D., Mariotti, A., & Soubies, F. (1999). Late Holocene phytolith and carbon-isotope record from a Latosol at Salitre, South-central Brazil. Quaternary Research, 51(2), 187-194. doi: https://doi.org/10.1006/qres.1998.2027
https://doi.org/10.1006/qres.1998.2027...
; Calegari et al., 2013Calegari, M. R., Madella, M., Vidal-Torrado, P., Pessenda, L. C. R., & Marques, F. A. (2013). Combining phytoliths and δ13C matter in Holocene palaeoenvironmental studies of tropical soils: an example of an Oxisol in Brazil. Quaternary International, 287, 47-55. doi: https://doi.org/10.1016/j.quaint.2011.11.012
https://doi.org/10.1016/j.quaint.2011.11...
; Fishkis et al., 2010Fishkis, O., Ingwersen, J., Lamers, M., Denysenko, D., & Streck, T. (2010). Phytolith transport in soil: a field study using flourescent labelling. Geoderma, 157(1-2), 27-36. doi: https://doi.org/10.1016/j.geoderma.2010.03.012
https://doi.org/10.1016/j.geoderma.2010....
) têm mostrado como a bioturbação e alixiviação são processos comuns que contribuem ao movimento vertical de fitólitos em solos naturais. Nesta situação (Alexandre et al. 1999Alexandre, A., Meunier, J.-D., Mariotti, A., & Soubies, F. (1999). Late Holocene phytolith and carbon-isotope record from a Latosol at Salitre, South-central Brazil. Quaternary Research, 51(2), 187-194. doi: https://doi.org/10.1006/qres.1998.2027
https://doi.org/10.1006/qres.1998.2027...
, p. 191), uma amostra de fitólitos seria resultado de uma mistura entre partículas mais jovens e outras mais antigas; porém, como a média da idade dos fitólitos é maior de acordo com o aumento da profundidade, eles ainda conseguiriam indicar mudanças na vegetação ao longo do tempo. No nosso caso, as terras pretas e o próprio aterro identificado na unidade N10049 E9956, no Teotônio, são construções humanas e, mesmo com certa mistura pós-deposicional dos solos/fitólitos, a integridade estratigráfica dos mesmos está apoiada pelo fato de que as maiores mudanças identificadas no registro correspondem às camadas definidas arqueologicamente. Para explorar melhor essa questão, teríamos que ampliar as análises de concentração nos demais níveis do perfil.
As primeiras três camadas (VI B1 a B3), entre 370-130 cm abaixo da superfície, estão localizadas onde foram encontradas as cerâmicas P-A misturadas com material que possivelmente havia sido redepositado por grupos posteriores que realizaram obras de terraplenagem (Figura 1, Kater et al., 2020Kater, T., Almeida, F. Almeida, F. O, Mongeló, G. Z.; Watling, J.; Neves, E. G. (2020). Variabilidade estratigráfica e espacial dos contextos cerâmicos no sítio Teotônio. Revista de Arqueologia, 33(1), 198-220.). Os fitólitos, na fração de silte dessas camadas (Figura 3), foram dominados por morfotipos de plantas lenhosas (globular granulates) e por palmeiras (Arecaceae), as últimas sendo mais abundantes e aumentando em frequência de baixo para cima do perfil. Essas camadas têm as frequências mais altas (> 15%) de gramíneas (Poaceae) do perfil inteiro, as quais foram representadas, em ordem descendente, pelas subfamílias Chloridoideae, Panicoideae, Bambusoideae e Arundinoideae. Na fração de areia (Figura 4), dois morfotipos potencialmente pertencentes à família Marantaceae e/ou a outras herbáceas monocotiledôneas (cf.Chen & Smith, 2013Chen, S. T., & Smith, S. Y. (2013). Phytolith variability in Zingiberales: a tool for the reconstruction of past tropical vegetation. Palaeogeography, Palaeoclimatology, Palaeoecology, 370, 1-12. doi: 10.1016/j.palaeo.2012.10.026
https://doi.org/10.1016/j.palaeo.2012.10...
) (Figuras 2K e 2L) aparecem de repente em grandes quantidades, junto com outros morfotipos diagnósticos em relação às Asteraceae, sementes de Marantaceae e bulliforms (silicificações de células motoras) de gramíneas – todas sendo plantas indicativas de uma vegetação significativamente aberta. Finalmente, quase todas as amostras analisadas destas camadas continham fitólitos de abóbora, ao passo que fitólitos dos cultivos de mandioca, araruta (Maranta arundinacea) (Figura 2M) e, possivelmente, ariá (Calathea allouia, cf.Chandler-Ezell et al., 2006Chandler-Ezell, K., Pearsall, D. M., & Zeidler, J. A. (2006). Root and tuber phytoliths and starch grains document manioc (Manihot escuelenta), arrowroot (Maranta aruniacea), and llerén (Calathea sp.) at the Real Alto site, Ecuador. Economic Botany, 60(2), 103-120.) (Figura 2N) foram recuperados em quantidades menores. A mandioca foi encontrada na amostra basal da unidade (360-370 cm).
Os fitólitos da camada VI C, considerada como uma camada de transição, apresentaram uma pequena ruptura. Enquanto as frequências de palmeiras e os tipos arbóreos ficaram iguais aos níveis inferiores, os fitólitos de ervas (gramíneas, Asteraceae e Marantaceae) diminuíram substancialmente neste nível. Fitólitos de milho (Figura 2O) aparecem aqui pela primeira vez. Uma das lentes de argila avermelhada, visíveis nesta camada, também foi analisada e apresentou um conjunto de fitólitos semelhante à camada ao redor.
A camada VI D, caraterizada pela TPA mais escura do sítio e pela presença de abundantes cerâmicas JM orientadas horizontalmente, foi considerada uma antiga superfície. Aqui, entre 80-50 cm, foram encontradas frequências consideráveis de fitólitos de milho (1-3%) e alguns fitólitos de cf. ariá, enquanto a abóbora esteve ausente. Fitólitos de palmeiras continuaram frequentes nesta camada, formando um pico iniciado ao final da camada VI B3 e que termina antes da camada E. Os gêneros Bactris/Astrocaryum spp. contribuíram aproximadamente com metade de todos os fitólitos de palmeiras encontrados.
A camada VI E, que consiste em TPA com cerâmicas JT, apresenta uma outra ruptura em termos dos fitólitos. Fitólitos de plantas lenhosas dobram em frequência nesta camada, ao custo das palmeiras. A amostra de base desta camada (50-60 cm) ainda continha fitólitos de milho, junto com um fitólito de mandioca, mas o milho depois desaparece entre 0-40 cm. A porcentagem elevada de plantas lenhosas continua até a camada VII, quando palmeiras e gramíneas (subfamília Panicoideae) aumentam novamente, representando as perturbações modernas.
SANTA PAULA
Os resultados da análise estratigráfica da unidade N1000 E1001 de Santa Paula estão apresentados nas Figuras 5 e 6.
Porcentagens de fitólitos da fração silt (0-53 µm) encontrados no Montículo 1 de Santa Paula.
Contagens absolutas de fitólitos da fração arenosa (53-250 µm) encontrados no Montículo 1 de Santa Paula.
As primeiras duas camadas analisadas, a I, estéril, e a II, são interpretadas como pertencentes a ocupações anteriores à construção do montículo e apresentaram fitólitos muito distintos dos níveis superiores, sendo dominadas por fitólitos de plantas lenhosas (globular granulates) e gramíneas. Nos fitólitos de gramíneas, foram especialmente frequentes os bulliforms e tipos da subfamília Bambusoideae.
Já nas camadas III (uma camada fina de material P-A misturado a outros contemporâneos e/ou mais antigos) e IV (camada construtiva da base do montículo, com cerâmicas BC misturadas às mais antigas), começa a aparecer um conjunto de espécies diferentes, quando as frequências de bulliforms diminuem e as de palmeiras aumentam de < 5% para 60%. Essa mesma tendência continua através das camadas V e VI (BC), onde as palmeiras atingem mais de 70% do total de fitólitos. Destaca-se, na camada VI, um aumento em fitólitos de Bactris/Astrocaryum spp., que contribuíram para aproximadamente metade do total das palmeiras.
Fitólitos de milho foram encontrados ao lado de cf. ariá em duas amostras entre 30-60 cm da camada VI; também foram encontrados sozinhos na amostra entre 130-140 cm, na base da camada V. Esses níveis onde o milho foi encontrado também têm as maiores concentrações de cerâmicas BC.
Na camada VII e na amostra superior da camada VI, na fração arenosa (Figura 6), aparecem os mesmos dois fitólitos não identificados que foram encontrados nas camadas VI B2 e B3 no Teotônio, possivelmente pertencentes à família Marantaceae ou a uma parente próxima. O seu aumento até o Horizonte A (0-10 cm) acontece junto com fitólitos das sementes da Marantaceae e da família Asteraceae, e sugere que esse sinal vem de atividades modernas, quando o terreno fazia parte de um sítio de moradores ribeirinhos.
DISCUSSÃO E INTERPRETAÇÃO
O USO DAS PLANTAS E SEUS CONTEXTOS DE DESCARTE
Os resultados obtidos neste estudo permitem avançar nas interpretações a respeito da relação das populações pré-coloniais com as plantas, na região do alto rio Madeira, e formular novas questões sobre os sistemas de manejo de plantas durante os últimos 3.000 anos na região.
TEOTÔNIO
Como apontamos anteriormente, ainda não sabemos qual foi a natureza da interação entre os produtores de cerâmica P-A e as populações da fase Massangana, quando aqueles chegaram na Cachoeira do Teotônio. Destaca-se, primeiro, a ubiquidade de amido de milho encontrado em quase todos os fragmentos dos vasos P-A analisados. Apesar das datas antigas para o milho no sudoeste amazônico (Brugger et al., 2016Brugger, S. O., Gobet, E., Van Leeuwen, J. F. N., Ledru, M. P., Colombaroli, D., Van Der Knaap, W. O., . . . Tinner, W. (2016). Long-term man-environment interactions in the Bolivian Amazon: 8000 years of vegetation dynamics. Quaternary Science Reviews, 132, 114-128. doi: 10.1016/j.quascirev.2015.11.001
https://doi.org/10.1016/j.quascirev.2015...
; Kistler et al., 2018Kistler, L., Maezumi, S. Y., Gregorio de Souza, J., Przelomska, N. A., Malaquias Costa, F., Smith, O., Loiselle, H., . . . Allaby, R. G. (2018). Multiproxy evidence highlights a complex evolutionary legacy of maize in South America. Science, 362(6420), 1309-1313. doi: http://doi.org/10.1126/science.aav0207
https://doi.org/10.1126/science.aav0207...
), ele não foi encontrado nos contextos pré-ceramistas do Teotônio (Watling et al., 2018Watling, J., Shock, M. P., Mongeló, G. Z., Almeida, F. O., Kater, T., De Oliveira, P. E., & Neves, E. G. (2018). Direct archaeological evidence for Southwestern Amazonia as an early plant domestication and food production centre. PLos ONE, 13(7), 1-28. doi: 10.1371/journal.pone.0199868
https://doi.org/10.1371/journal.pone.019...
), gerando a pergunta se esse cultivo foi introduzido à região com as ocupações P-A – para a qual ainda não temos uma resposta. Kater et al. (2020)Kater, T. (2020). A temporalidade das ocupações ceramistas no sítio Teotônio. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, 15(2), e20190078. doi: 10.1590/2178-2547-BGOELDI-2019-0078
https://doi.org/10.1590/2178-2547-BGOELD...
mostraram como as formas dos vasilhames P-A no Teotônio têm uma forte tendência a serem vasos com pequena capacidade volumétrica, o que indicaria formas ligadas ao consumo, mas não ao processamento de alimentos. Os autores relacionam esse padrão, ausente nos outros sítios na área, com a própria forma de uso do sítio por essas populações, possivelmente engajadas em eventos ou rituais de consumo de bebidas no lado da cachoeira (Almeida & Kater, 2017Almeida, F. O., & Kater, T. (2017). As cachoeiras como bolsões de histórias dos grupos indígenas das terras baixas sul-americanas. Revista Brasileira de História, 37(75), 39-67. doi: 10.1590/1806-93472017v37n75-02a
https://doi.org/10.1590/1806-93472017v37...
; Kater et al., 2020Kater, T., Almeida, F. Almeida, F. O, Mongeló, G. Z.; Watling, J.; Neves, E. G. (2020). Variabilidade estratigráfica e espacial dos contextos cerâmicos no sítio Teotônio. Revista de Arqueologia, 33(1), 198-220.). Os danos presentes nos grãos de amido encontrados nos vasos indicam que as plantas sofreram uma variedade de processos mecânicos (e.g. moagem) e de aquecimento (tostado e fervido). Apesar de danos diagnósticos de fermentação (Henry et al., 2009Henry, A. G., Hudson, H. F., & Piperno, D. R. (2009). Changes in starch grain morphologies from cooking. Journal of Archaeological Science, 36(3), 915-922. doi: 10.1016/j.jas.2008.11.008
https://doi.org/10.1016/j.jas.2008.11.00...
; Vinton et al., 2009Vinton, S. D., Perry, L., Reinhard, K. J., Santoro, C. M., & Teixeira-Santos, I. (2009). Impact of empire expansion on household diet: the Inka in northern Chile’s Atacama Desert. PLoS ONE, 4(11), 1-5. doi: 10.1371/journal.pone.0008069
https://doi.org/10.1371/journal.pone.000...
) não terem sido encontrados, os dados não excluem a possibilidade de que estamos diante do processo de produção de bebidas fermentadas de milho.
Devido à provável situação de deposição secundária das camadas VI B1 a B3 por ocupações posteriores da tradição P-A, os fitólitos dos solos podem ter vindo de uma variedade de contextos que se formaram entre aproximadamente 3.000 e 1.000 cal. A.P. Entre o material Pocó-Açutuba, nessas camadas encontram-se cerâmicas da tradição Barrancoide/Borda Incisa (presentes em Santa Paula ca. 1.600 cal. A.P.) e do conjunto Morro dos Macacos (presente regionalmente ca. 1.800 cal. A.P.) (Kater et al., 2020Kater, T. (2020). A temporalidade das ocupações ceramistas no sítio Teotônio. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, 15(2), e20190078. doi: 10.1590/2178-2547-BGOELDI-2019-0078
https://doi.org/10.1590/2178-2547-BGOELD...
). Essas camadas são ricas em fitólitos de cultivos, e a sua presença junto com altos teores de gramíneas e outras ervas sugere que essas TPA se formaram originalmente em um ambiente de vegetação aberta, provavelmente em clareiras próximas às casas, em áreas usadas como jardins e descarte de lixo (sensuSchmidt et al., 2014Schmidt, M. J., Py-Daniel, A. R., Morães, C. P., Valle, R. B. M., Caromano, C. F., Texeira, W. G., . . . Heckenberger, M. J. (2014). Dark earths and the human built landscape in Amazonia: a widespread pattern of anthrosol formation. Journal of Archaeological Science, 42, 152-165. doi: 10.1016/j.jas.2013.11.002
https://doi.org/10.1016/j.jas.2013.11.00...
). A alta frequência de fitólitos de palmeiras apoia esse cenário. Amplamente usadas no consumo e na construção de casas, essas plantas provavelmente foram manejadas e descartadas nesses mesmos contextos.
No sítio Teotônio, a ocupação marcada pelas cerâmicas da tradição JM é representada por uma mudança abrupta na estratigrafia da unidade analisada. Com TPA de cor preta, as cerâmicas são abundantes e organizadas horizontalmente, sugerindo tratar-se de uma antiga superfície de ocupação (Figura 1). Na camada VI D e na camada de transição abaixo (VI C), expressivas quantidades de fitólitos de milho, junto com cf. ariá, apoiam a interpretação de que aqui havia uma superfície ativa de deposição. As cerâmicas JM também continham grãos de amido de milho, alguns com danos físicos iguais àqueles observados no material P-A. Os vasos desta tradição são de várias morfologias e capacidades (até 80 litros) e apresentam fuligem resultante da exposição ao fogo (Kater, 2018Kater, T. (2018). O sítio Teotônio e as reminiscências de uma longa história indígena no alto Rio Madeira (Mestrado em Arqueologia). Universidade Federal de Sergipe, Sergipe, Aracajú.; Kater et al., 2020Kater, T., Almeida, F. Almeida, F. O, Mongeló, G. Z.; Watling, J.; Neves, E. G. (2020). Variabilidade estratigráfica e espacial dos contextos cerâmicos no sítio Teotônio. Revista de Arqueologia, 33(1), 198-220.), sugerindo que foram usados utilitariamente para a preparação de alimentos. Amidos de abóbora e outros taxa não identificados também estavam presentes nas cerâmicas JM, tanto como nas cerâmicas P-A. Porém, contrastando com as camadas inferiores, fitólitos de abóbora desaparecem completamente dos solos deste contexto.
No trabalho de Alves (2017)Alves, D. T. (2017). Dark earth plant management in the lower Tapajos (Doutorado em Arqueologia). Universidade de Exeter, Exeter., que analisou fitólitos de TPA na região de Santarém e Belterra, foram encontrados fitólitos das espigas de milho apenas em contextos domésticos, junto a TPA mais escura, enquanto a abóbora esteve presente ao longo das estratigrafias inteiras dos perfis analisados, os quais incluíram camadas de TPA mais amarronzadas (Alves, 2017Alves, D. T. (2017). Dark earth plant management in the lower Tapajos (Doutorado em Arqueologia). Universidade de Exeter, Exeter.). É possível que o aumento repentino de milho nas camadas Jamari também seja um produto de área de atividade, se tivermos diante de um contexto doméstico. Porém, a quantidade de cerâmicas com a presença de fuligem dentro de fraturas e quebras na camada VI D sugere que este contexto era uma área de lixeira onde restos orgânicos e inorgânicos foram jogados e queimados posteriormente ao descarte (Kater, 2018Kater, T. (2018). O sítio Teotônio e as reminiscências de uma longa história indígena no alto Rio Madeira (Mestrado em Arqueologia). Universidade Federal de Sergipe, Sergipe, Aracajú.; Kater et al., 2020Kater, T., Almeida, F. Almeida, F. O, Mongeló, G. Z.; Watling, J.; Neves, E. G. (2020). Variabilidade estratigráfica e espacial dos contextos cerâmicos no sítio Teotônio. Revista de Arqueologia, 33(1), 198-220.). É interessante como a concentração de fitólitos no nível 70-80 cm (da camada VI D) é semelhante àquela das camadas VI B1 a B3, apoiando a interpretação de que as últimas originalmente se formaram também em contextos de lixeiras, antes de serem redepositadas. A concentração de fitólitos de espigas de milho somente na camada VI D, portanto, sugere descarte e, subsequentemente, consumo mais alto dessa espécie nas ocupações JM do que nas anteriores.
As altas frequências de fitólitos de palmeiras também sugerem intenso descarte dessas espécies nas Camadas Jamari, algo que, na realidade, é uma continuação do padrão das ocupações prévias (Figura 3). São nestas camadas, e na camada VI B3, que fitólitos dos gêneros Bactris/Astrocaryum spp. aparecem em teores mais altos, constituindo metade do total dos fitólitos de palmeiras. Esses gêneros contêm as espécies pupunha (Bactris gasipaes) e tucumã (Astrocaryum vulgare). Embora saibamos que a pupunha foi domesticada na região sudoeste da Amazônia e se dispersou seguindo o rio Madeira (Clement et al., 2017Clement, C. R., Cristo-Araújo, M., Coppens d’Eeckenbrugge, G., Reis, V. M., Lehnebach, R., & Picanço-Rodrigues, D. (2017). Origin and dispersal of domesticated peach palm. Frontiers in Ecology and Evolution, 5, 1-19. doi: 10.3389/fevo.2017.00148
https://doi.org/10.3389/fevo.2017.00148...
), o tucumã talvez seja a palmeira mais presente nos jardins atuais dos moradores na vila Teotônio (observação de J. Watling, em 2018). Futuros estudos taxonômicos dos vestígios macrobotânicos nestes contextos têm grande potencial de elucidar de quais dessas espécies vieram os fitólitos.
Finalmente, tratamos os dados das ocupações pertencentes à subtradição Jatuarana, manifestação local da tradição Polícroma da Amazônia, tradição pan-amazônica, que é a última representada na sequência do alto Madeira antes dos europeus (Almeida, 2013Almeida, F. O. (2013). A Tradição Polícroma no alto rio Madeira (Doutorado em Arqueologia). Universidade de São Paulo, São Paulo.; Almeida & Moraes, 2016Almeida, F. O., & Moraes, C. P. (2016). A cerâmica polícroma do rio Madeira. In C. Barreto, H. P. Lima, & C. Jaimes Betancourt (Orgs.), Cerâmicas arqueológicas da Amazônia: rumo a uma nova síntese (pp. 402-413). Belém: IPHAN.). Enquanto a idade dos materiais analisadas aqui está incerta, a cerâmica JT aparece na região do alto Madeira aproximadamente há ca. 1.300 cal. A.P. Os fitólitos da camada VI E, que apresenta TPA tão escura quanto nas camadas JM, parecem indicar uma ruptura significativa na maneira segundo a qual as pessoas estavam vivendo neste lugar. A diminuição de Bactris/Astrocaryum spp. e o aumento drástico de plantas lenhosas sugerem que houve maior concentração dos últimos crescendo localmente e/ou sendo descartados no período JT. A baixa resolução taxonômica de fitólitos produzidos por árvores, em geral, não nos permite dizer se estes vestígios vieram de espécies úteis. Dos oito fragmentos de cerâmica analisados, apenas um forneceu significantes quantidades de grãos de amido. A maioria desses grãos, porém, não correspondeu a nenhum dos morfotipos já publicados, enquanto três foram tentativamente atribuídos à mandioca. Há duas hipóteses em relação à ausência de amido nas cerâmicas JT: i) a alta fragmentação deste material fez com que as análises fossem comprometidas pelo tamanho da superfície disponível para a extração de microvestígios, ou ii) essas cerâmicas foram pouco utilizadas antes de serem descartadas.
O achado de dois artefatos de quartzo recuperados do mesmo contexto, cheios de grãos de amido de mandioca, porém, é um forte indício dos famosos dentes de ralador de mandioca e, portanto, de mandioca brava. Do mesmo modo que Perry (2002)Perry, L. (2002). Starch analyses reveal multiple functions of quartz “manioc” grater flakes from the Orinoco Basin, Venezuela. Interciencia, 27(11), 635-639. encontrou grãos de amido de uma variedade de cultivos em dentes de ralador do baixo Orinoco, os artefatos aqui analisados também foram utilizados para processar espécies diferentes, como batata doce (a segunda espécie mais abundante) e, possivelmente, cará e feijão. Apenas um dos três líticos da ocupação JM continha grãos de amido e nenhum era oriundo de plantas tuberosas (um de milho e dois não identificados).
Ainda que sejam necessárias mais análises dos líticos das demais ocupações ceramistas, podemos levantar a hipótese, a ser testada futuramente, de que os portadores de cerâmica JT trouxeram consigo, para o Teotônio, um novo conjunto de recursos vegetais, dando maior ênfase às plantas tuberosas e uma ênfase menor ao milho, se compararmos com a ocupação JM.
SANTA PAULA
O sítio Santa Paula, embora seja diferente do Teotônio em quase todos os aspectos, apresenta semelhanças em relação ao registro microbotânico, as quais merecem atenção.
O montículo de TPA analisado aqui foi construído em torno de 1.600 cal. A.P. O registro das camadas subjacentes sugere a presença de uma paisagem aberta, dominada por gramíneas, que foi visitada frequentemente pelos produtores de cerâmicas P-A e outras, os quais deixaram carvão e cerâmicas na superfície do sítio. A camada V, que representa uma camada de ocupação e/ou construtiva do montículo (Figura 1), consiste em TPA escura, igual à fase JM no Teotônio, só que está repleta de feições que sugerem a presença de casas, sendo, então, uma mistura de material secundário e in situ (Bespalez et al., 2020Bespalez, E., Zuse, S., Pessoa, C., Venere, P., & Santi, J. (2020). Arqueologia no sítio Santa Paula, alto Madeira, Porto Velho, Roraima, Brasil. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, 15(2), e20190076. doi: 10.1590/2178-2547-BGOELDI-2019-0075
https://doi.org/10.1590/2178-2547-BGOELD...
). Nessa camada, não há vestígios de plantas comestíveis (com exceção do milho, na base da camada), diferentemente do que foi encontrado no Teotônio, nas camadas construtivas do possível aterro.
Na camada VI, onde a TPA é mais escura, a presença de fitólitos de milho e cf. ariá sugere que os depósitos entre 30-60 cm se tratam de um local onde havia uma antiga superfície com atividades domésticas ou uma área de descarte. Embora a recuperação de grãos de amido das cerâmicas BC analisadas tenha sido baixa, a espécie mais representada foi o milho. Adicionalmente, a presença de fitólitos de mandioca em dois dos artefatos é notável. Diante disso, questiona-se: por que não foi encontrado amido de mandioca nos resíduos dos mesmos artefatos? Há pelo menos duas explicações alternativas: i) o processamento intensivo da mandioca fez com que nenhum grão de amido sobrevivesse ou ii) estamos diante de um produto feito de uma outra parte da planta que não seja o tubérculo, como a folha (e.g. maniçoba), que não contém amido diagnóstico, mas que produz fitólitos (Chandler-Ezell et al., 2006Chandler-Ezell, K., Pearsall, D. M., & Zeidler, J. A. (2006). Root and tuber phytoliths and starch grains document manioc (Manihot escuelenta), arrowroot (Maranta aruniacea), and llerén (Calathea sp.) at the Real Alto site, Ecuador. Economic Botany, 60(2), 103-120.). Além da mandioca e do milho, podemos incluir o cará ao conjunto de cultivos consumidos neste sítio.
Uma das coisas mais interessantes no registro microbotânico de Santa Paula é a alta frequência de fitólitos de palmeiras, superando as contagens das camadas VI B3, VI C e VI D no Teotônio (Figuras 1 e 3), os quais, novamente, estão dominados por espécie(s) dentro dos gêneros Bactris/Astrocaryum spp. Neste caso, sendo um contexto doméstico, e não uma lixeira, poderíamos hipotetizar que estes fitólitos vieram do material construtivo usado nas casas, e que os mesmos fitólitos encontrados nas TPA das lixeiras no Teotônio também tiveram a mesma origem.
É intrigante que o aumento de fitólitos de Bactris/Astrocaryum spp., relativos a outras espécies de palmeiras, é um fenômeno registrado em sítios muitos distantes uns dos outros na bacia amazônica, sempre associado com ocupações em TPA escura. Como já destacado por Alves (2017)Alves, D. T. (2017). Dark earth plant management in the lower Tapajos (Doutorado em Arqueologia). Universidade de Exeter, Exeter., um aumento na recuperação de Bactris/Astrocaryum spp. é visto tanto nos sítios na região de Santarém quanto nas camadas superiores do sítio Hatahara, perto de Manaus (Bozarth et al., 2009Bozarth, S. R., Price, K., Woods, W. I., Neves, E. G., & Rebellato, L. (2009). Phytoliths and Terra Preta: the Hatahara site example. In W. I. Woods, W. G. Texeira, J. Lehmann, C. Steiner, A. M. G. A. Winklerprins & L. Rebellato (Orgs.), Amazonian Dark Earths: Will Sombroek’s Vision (pp. 85-98). New York: Springer.; Cascon, 2010Cascon, L. M. (2010). Alimentação na floresta tropical: um estudo de caso no sítio Hatahara, Amazônia Central, com base em microvestígios botânicos (Mestrado em Arqueologia). Universidade Federal do Rio de Janeiro, Rio de Janeiro.) – sendo esses contextos regionais datados aproximadamente entre 1.500 e 1.000 cal. A.P. Quais espécies estamos vendo e para quais fins foram utilizadas? Estudos macrobotânicos serão essenciais para responder a essas perguntas no futuro.
A camada com material cerâmico JT, que está presente entre 0 e 40 cm, misturado ao BC, apresenta fitólitos que parecem vir de atividades modernas, o que não permite fazer muitas observações. A ausência de fitólitos de milho nos superiores a 30 cm é, porém, interessante, sob a luz dos achados no Teotônio.
RESUMO DOS RESULTADOS
Com base nos dados apresentados, pode-se voltar às perguntas originais levantadas na Introdução:
-
Podemos sugerir mudanças no uso de plantas (como em quais das plantas foram cultivadas e o contextos de consumo e de uso) ao longo da sequência de ocupações ceramistas no Teotônio?
A partir dos dados apresentados aqui, levantamos algumas hipóteses preliminares que indicam tendências nos dados de um registro extremamente complexo: 1) os contextos com cerâmicas P-A (3.000 a 1.500 cal. A.P.) são os mais antigos associados à presença do milho, de maneira significativa, na calha do alto rio Madeira; 2) nos mesmos contextos, há evidências de utilização do milho para fazer bebidas, que podem ter sido consumidas coletivamente em festas (sensuKater et al., 2020Kater, T., Almeida, F. Almeida, F. O, Mongeló, G. Z.; Watling, J.; Neves, E. G. (2020). Variabilidade estratigráfica e espacial dos contextos cerâmicos no sítio Teotônio. Revista de Arqueologia, 33(1), 198-220.); 3) entre ca. 3.000-1.500 cal. A.P., as populações no alto Madeira praticavam um sistema de manejo agroflorestal focado em múltiplos cultivos, como abóbora, milho, araruta e ariá. Certamente, este sistema incluiu inúmeras espécies de palmeiras e árvores cultivadas, mas somente as palmeiras deixaram seus sinais no registro microbotânico; 4) as ocupações associadas a cerâmicas Jamari têm evidências de economias fortemente baseadas no cultivo de milho, embora ainda se aproveitando dos recursos agroflorestais presentes no sítio e em seus arredores; e 5) ocupações Jatuarana têm evidências de dietas mais orientadas ao cultivo de tubérculos como a mandioca, dentro de um sistema de policultura agroflorestal.
-
Como o registro arqueobotânico de Santa Paula se relaciona com aquele do Teotônio?
O registro do Santa Paula, sendo particular e de mais curta duração, dificulta comparações cronológicas diacrônicas no nível possibilitado pelo Teotônio. Podemos dizer que os contextos associados a cerâmicas Barrancoide/Borda Incisa têm presença de milho, e possivelmente ariá, com outros alimentos ainda sendo desconhecidos. A alta frequência de Bactris/Astrocaryum spp. no registro é, porém, igual ao que se registra no Teotônio, sendo um fenômeno que ocorre em outros sítios de TPA muito distantes, no mesmo período.
-
O que os vestígios botânicos podem nos dizer sobre a maneira de ocupar a paisagem, incluindo a própria formação dos sítios?
A interpretação de dados de tecnologia cerâmica, estratigrafia e contextos de preservação, juntamente com dados microbotânicos, tem permitido um olhar mais informado em relação ao registro arqueológico nos dois sítios analisados, principalmente no Teotônio, permitindo que se identifique quais elementos se modificaram e quais permaneceram na história de 3.000 anos de cultivo e manejo de recursos vegetais na região. Ademais, foi possível sugerir uma possível origem das camadas redepositadas do ‘aterro’ através dos tipos de fitólitos encontrados, assim como averiguar possíveis paleossuperfícies na estratigrafia.
CONSIDERAÇÕES FINAIS
A arqueologia amazônica tem avançado muito desde as teorias de determinismo agrícola e ambiental (Meggers, 1954Meggers, B. J. (1954). Environmental limitations on the development of culture. American Anthropologist, 56, 801-824.; Lathrap, 1970Lathrap, D. W. (1970). The Upper Amazon. London: Thames and Hudson.; Roosevelt, 1980Roosevelt, A. C. (1980). Parmana: prehistoric maize and manioc subsistence along the Amazon and Orinoco. New York: Academic Press.), para incorporar teorias de pluralismo, abundância e diversidade dentro de sistemas de manejo (Balée, 2010Balée, W. (2010). Contingent diversity on anthropic landscapes. Diversity, 2(2), 163-181. doi: 10.3390/d2020163
https://doi.org/10.3390/d2020163...
; Moraes, 2015Moraes, C. P. (2015). O determinismo agrícola na arqueologia amazônica. Estudos Avançandos, 29(83), 25-43. doi: http://dx.doi.org/10.1590/S0103-40142015000100004
https://doi.org/10.1590/S0103-4014201500...
; Fausto & Neves, 2018Fausto, C., & Neves, E. G. (2018). Was there ever a Neolithic in the Neotropics? Plant familiarisation and biodiversity in the Amazon. Antiquity, 92(366), 1604-1618. doi: 10.15184/aqy.2018.157
https://doi.org/10.15184/aqy.2018.157...
; Levis et al., 2018Levis, C., Flores, B. M., Moreira, P. A., Luize, B. G., Alves, R. P., Franco-Moraes, J., . . . Clement, C. R. (2018). How people domesticated Amazonian forests. Frontiers in Ecology and Evolution, 5(171). doi: 10.3389/fevo.2017.00171
https://doi.org/10.3389/fevo.2017.00171...
). Entretanto, é importante que continuemos a estudar essas práticas para não ficarmos presos nas mesmas armadilhas essencialistas que guiaram o pensamento até os fins dos anos 80.
Este estudo se soma a um corpo crescente de trabalhos microarqueobotânicos feitos nas TPA amazônicas do Holoceno tardio (Bozarth et al., 2009Bozarth, S. R., Price, K., Woods, W. I., Neves, E. G., & Rebellato, L. (2009). Phytoliths and Terra Preta: the Hatahara site example. In W. I. Woods, W. G. Texeira, J. Lehmann, C. Steiner, A. M. G. A. Winklerprins & L. Rebellato (Orgs.), Amazonian Dark Earths: Will Sombroek’s Vision (pp. 85-98). New York: Springer.; Cascon, 2010Cascon, L. M. (2010). Alimentação na floresta tropical: um estudo de caso no sítio Hatahara, Amazônia Central, com base em microvestígios botânicos (Mestrado em Arqueologia). Universidade Federal do Rio de Janeiro, Rio de Janeiro.; Alves, 2017Alves, D. T. (2017). Dark earth plant management in the lower Tapajos (Doutorado em Arqueologia). Universidade de Exeter, Exeter.; Maezumi et al., 2018Maezumi, S. Y., Alves, D., Robinson, M., Gregorio de Souza, J., Levis, C., Barnett, R. L., . . . Iriarte, J. (2018). The legacy of 4,500 years of polyculture agroforestry in the eastern Amazon. Nature Plants, 4(8), 540-547. doi: 10.1038/s41477-018-0205-y
https://doi.org/10.1038/s41477-018-0205-...
; Watling et al., 2018Watling, J., Shock, M. P., Mongeló, G. Z., Almeida, F. O., Kater, T., De Oliveira, P. E., & Neves, E. G. (2018). Direct archaeological evidence for Southwestern Amazonia as an early plant domestication and food production centre. PLos ONE, 13(7), 1-28. doi: 10.1371/journal.pone.0199868
https://doi.org/10.1371/journal.pone.019...
). Embora seja evidente que as populações que criaram esses solos, a partir de 3.000 anos atrás, engajavam-se na policultura, incluindo o manejo de muitas plantas e árvores silvestres ou na ‘escala de domesticação’ (Clement et al., 2015Clement, C. R., Denevan, W. M., Heckenberger, M. J., Junqueira, A. B., Neves, E. G., Texeira, W. G., & Woods, W. I. (2015). The domestication of Amazonia before European conquest. Proceedings of the Royal Society B, 282(1812), 1-9. doi: 10.1098/rspb.2015.0813
https://doi.org/10.1098/rspb.2015.0813...
; Maezumi et al., 2018Maezumi, S. Y., Alves, D., Robinson, M., Gregorio de Souza, J., Levis, C., Barnett, R. L., . . . Iriarte, J. (2018). The legacy of 4,500 years of polyculture agroforestry in the eastern Amazon. Nature Plants, 4(8), 540-547. doi: 10.1038/s41477-018-0205-y
https://doi.org/10.1038/s41477-018-0205-...
), é possível que em algumas regiões e em alguns momentos houvesse uma ênfase maior em uma só planta. Nos Llanos de Mojos, na Bolívia, entre aproximadamente 1.500 e 600 cal. A.P., estudos arqueobotânicos mostram grande diversidade de cultivos (Dickau et al., 2012Dickau, R., Bruno, M. C., Iriarte, J., Prümers, H., Jaimes Betancourt, C., Holst, I., & Mayle, F. E. (2012). Diversity of cultivars and other plant resources used at habitation sites in the Llanos de Mojos, Beni, Bolivia: evidence from macrobotanical remains, starch grains, and phytoliths. Journal of Archaeological Science, 39(2), 357-370. doi: 10.1016/j.jas.2011.09.021
https://doi.org/10.1016/j.jas.2011.09.02...
), porém, a grande maioria dos vestígios nos assentamentos e nos campos elevados é de milho (Iriarte & Dickau, 2012Iriarte, J., & Dickau, R. (2012). Las culturas del maíz? Arquebotânica de las sociedades hidráulicas de las tierras bajas sudamericanas. Amazônica, 4, 30-58.; Bruno, 2015Bruno, M. C. (2015). Macrorestos botánicos de la Loma Mendoza. In H. Prümers (Ed.), Loma Mendoza: las excavaciones de los años 1999-2002 (pp. 285-296). La Paz: Plural Editores.). No alto rio Xingu, a cultura Xinguana parece ter cultivado a mandioca como alimento de base na alimentação, pelo menos nos últimos mil anos (Heckenberger, 1998Heckenberger, M. J. (1998). Manioc agriculture and sedentism in Amazonia: the Upper Xingu example. Issues in Brazilian Archaeology, 2, 633-648.). No alto rio Madeira, começa-se a perceber que o milho teve importância significativa há ca. 1.000 anos atrás, com as ocupações da tradição Jamari e Barrancoide/Borda Incisa.
Futuros estudos no alto rio Madeira poderão confirmar ou não as hipóteses apresentadas aqui, e deverão focar: i) no estudo do registro macrobotânico dos mesmos contextos, para acessar a gama de espécies frutíferas que estavam sendo manejadas ao lado das plantas domesticadas; ii) na ampliação dos estudos microbotânicos para incorporar mais artefatos líticos, buscando verificar a cronologia dos possíveis dentes de ralador na região; iii) na ampliação do número de intervenções nos sítios e fora dos sítios, para abranger diferentes contextos representativos; e – por último mas não menos importante – iv) na expansão de coleções de referência de plantas modernas, para poder identificar as dezenas de grãos de amido observados neste estudo, os quais não puderam ser taxonomicamente identificados.
-
Watling, J., Almeida, F., Kater, T., Zuse, S., Shock, M. P., Mongeló, G., Bespalez, E., Santi, J. R., & Neves, E. G. (2020)Watling, J., Almeida, F., Kater, T., Zuse, S., Shock, M. P., Mongeló, G., Bespalez, E., Santi, J. R., & Neves, E. G. (2020). Arqueobotânica das ocupações ceramistas na Cachoeira do Teotônio. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, 15(2), e20190075. doi: 10.1590/2178-2547-BGOELDI-2019-0075
https://doi.org/10.1590/2178-2547-BGOELD... . Arqueobotânica das ocupações ceramistas na Cachoeira do Teotônio. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, 15(2), e20190075. doi: 10.1590/2178-2547-BGOELDI-2019-0075
REFERÊNCIAS
- Alexandre, A., Meunier, J.-D., Mariotti, A., & Soubies, F. (1999). Late Holocene phytolith and carbon-isotope record from a Latosol at Salitre, South-central Brazil. Quaternary Research, 51(2), 187-194. doi: https://doi.org/10.1006/qres.1998.2027
» https://doi.org/10.1006/qres.1998.2027 - Almeida, F. O. (2013). A Tradição Polícroma no alto rio Madeira (Doutorado em Arqueologia). Universidade de São Paulo, São Paulo.
- Almeida, F. O., & Moraes, C. P. (2016). A cerâmica polícroma do rio Madeira. In C. Barreto, H. P. Lima, & C. Jaimes Betancourt (Orgs.), Cerâmicas arqueológicas da Amazônia: rumo a uma nova síntese (pp. 402-413). Belém: IPHAN.
- Almeida, F. O., & Kater, T. (2017). As cachoeiras como bolsões de histórias dos grupos indígenas das terras baixas sul-americanas. Revista Brasileira de História, 37(75), 39-67. doi: 10.1590/1806-93472017v37n75-02a
» https://doi.org/10.1590/1806-93472017v37n75-02a - Alves, D. T. (2017). Dark earth plant management in the lower Tapajos (Doutorado em Arqueologia). Universidade de Exeter, Exeter.
- Alves-Pereira, A., Clement, C. R., Picanço-Rodrigues, D., Veasey, E. A., Dequigiovanni, G., Ramos, S. L. F., . . . Zucchi, M. I. (2018). Patterns of nuclear and chloroplast genetic diversity and structure of manioc along major Brazilian Amazonian rivers. Annals of Botany, 121(4), 625-639. doi: 10.1093/aob/mcx190
» https://doi.org/10.1093/aob/mcx190 - Arroyo-Kalin, M. (2010). The Amazonian formative: crop domestication and anthropogenic soils. Diversity, 2(4), 473-504. doi: 10.3390/d2040473
» https://doi.org/10.3390/d2040473 - Babot, M. P. (2003). Starch grain damage as an indicator of food processing. In D. M. Hart & L. A. Wallis (Orgs.), Phytolith and starch research in the Australian-Pacific-Asian regions: the state of the art (pp. 69-81). Canberra: Pandanus Books.
- Balée, W. (2010). Contingent diversity on anthropic landscapes. Diversity, 2(2), 163-181. doi: 10.3390/d2020163
» https://doi.org/10.3390/d2020163 - Bespalez, E., Zuse, S., Pessoa, C., Venere, P., & Santi, J. (2020). Arqueologia no sítio Santa Paula, alto Madeira, Porto Velho, Roraima, Brasil. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, 15(2), e20190076. doi: 10.1590/2178-2547-BGOELDI-2019-0075
» https://doi.org/10.1590/2178-2547-BGOELDI-2019-0075 - Bozarth, S. R., Price, K., Woods, W. I., Neves, E. G., & Rebellato, L. (2009). Phytoliths and Terra Preta: the Hatahara site example. In W. I. Woods, W. G. Texeira, J. Lehmann, C. Steiner, A. M. G. A. Winklerprins & L. Rebellato (Orgs.), Amazonian Dark Earths: Will Sombroek’s Vision (pp. 85-98). New York: Springer.
- Brugger, S. O., Gobet, E., Van Leeuwen, J. F. N., Ledru, M. P., Colombaroli, D., Van Der Knaap, W. O., . . . Tinner, W. (2016). Long-term man-environment interactions in the Bolivian Amazon: 8000 years of vegetation dynamics. Quaternary Science Reviews, 132, 114-128. doi: 10.1016/j.quascirev.2015.11.001
» https://doi.org/10.1016/j.quascirev.2015.11.001 - Bruno, M. C. (2015). Macrorestos botánicos de la Loma Mendoza. In H. Prümers (Ed.), Loma Mendoza: las excavaciones de los años 1999-2002 (pp. 285-296). La Paz: Plural Editores.
- Calegari, M. R., Madella, M., Vidal-Torrado, P., Pessenda, L. C. R., & Marques, F. A. (2013). Combining phytoliths and δ13C matter in Holocene palaeoenvironmental studies of tropical soils: an example of an Oxisol in Brazil. Quaternary International, 287, 47-55. doi: https://doi.org/10.1016/j.quaint.2011.11.012
» https://doi.org/10.1016/j.quaint.2011.11.012 - Cascon, L. M. (2010). Alimentação na floresta tropical: um estudo de caso no sítio Hatahara, Amazônia Central, com base em microvestígios botânicos (Mestrado em Arqueologia). Universidade Federal do Rio de Janeiro, Rio de Janeiro.
- Chandler-Ezell, K., Pearsall, D. M., & Zeidler, J. A. (2006). Root and tuber phytoliths and starch grains document manioc (Manihot escuelenta), arrowroot (Maranta aruniacea), and llerén (Calathea sp.) at the Real Alto site, Ecuador. Economic Botany, 60(2), 103-120.
- Chen, S. T., & Smith, S. Y. (2013). Phytolith variability in Zingiberales: a tool for the reconstruction of past tropical vegetation. Palaeogeography, Palaeoclimatology, Palaeoecology, 370, 1-12. doi: 10.1016/j.palaeo.2012.10.026
» https://doi.org/10.1016/j.palaeo.2012.10.026 - Clement, C. R., Denevan, W. M., Heckenberger, M. J., Junqueira, A. B., Neves, E. G., Texeira, W. G., & Woods, W. I. (2015). The domestication of Amazonia before European conquest. Proceedings of the Royal Society B, 282(1812), 1-9. doi: 10.1098/rspb.2015.0813
» https://doi.org/10.1098/rspb.2015.0813 - Clement, C. R., Rodrigues, D. P., Alves-Pereira, A., Mühlen, G. S., Cristo-Araújo, M., Moreira, P. A., . . . Reis, V. M. (2016). Crop domestication in the upper Madeira River basin. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, 11(1), 193-205. doi: 10.1590/1981.81222016000100010
» https://doi.org/10.1590/1981.81222016000100010 - Clement, C. R., Cristo-Araújo, M., Coppens d’Eeckenbrugge, G., Reis, V. M., Lehnebach, R., & Picanço-Rodrigues, D. (2017). Origin and dispersal of domesticated peach palm. Frontiers in Ecology and Evolution, 5, 1-19. doi: 10.3389/fevo.2017.00148
» https://doi.org/10.3389/fevo.2017.00148 - Denevan, W. M. (2006). Pre-European forest cultivation in Amazonia. In W. Balée & C. L. Erickson (Orgs.), Time and complexity in historical ecology (pp. 153-164). New York: Columbia University Press.
- Dickau, R., Ranere, A. J., & Cooke, R. G. (2007). Starch grain evidence for the preceramic dispersals of maize and root crops into tropical dry and humid forests of Panama. Proceedings of the National Academy of Sciences, 104(9), 3651-3656. doi: 10.1073/pnas.0611605104
» https://doi.org/10.1073/pnas.0611605104 - Dickau, R., Bruno, M. C., Iriarte, J., Prümers, H., Jaimes Betancourt, C., Holst, I., & Mayle, F. E. (2012). Diversity of cultivars and other plant resources used at habitation sites in the Llanos de Mojos, Beni, Bolivia: evidence from macrobotanical remains, starch grains, and phytoliths. Journal of Archaeological Science, 39(2), 357-370. doi: 10.1016/j.jas.2011.09.021
» https://doi.org/10.1016/j.jas.2011.09.021 - Fausto, C., & Neves, E. G. (2018). Was there ever a Neolithic in the Neotropics? Plant familiarisation and biodiversity in the Amazon. Antiquity, 92(366), 1604-1618. doi: 10.15184/aqy.2018.157
» https://doi.org/10.15184/aqy.2018.157 - Fishkis, O., Ingwersen, J., Lamers, M., Denysenko, D., & Streck, T. (2010). Phytolith transport in soil: a field study using flourescent labelling. Geoderma, 157(1-2), 27-36. doi: https://doi.org/10.1016/j.geoderma.2010.03.012
» https://doi.org/10.1016/j.geoderma.2010.03.012 - Heckenberger, M. J. (1998). Manioc agriculture and sedentism in Amazonia: the Upper Xingu example. Issues in Brazilian Archaeology, 2, 633-648.
- Henry, A. G., Hudson, H. F., & Piperno, D. R. (2009). Changes in starch grain morphologies from cooking. Journal of Archaeological Science, 36(3), 915-922. doi: 10.1016/j.jas.2008.11.008
» https://doi.org/10.1016/j.jas.2008.11.008 - Hilbert, L., Neves, E. G., Pugliese, F., Whitney, B. S., Shock, M. P., Veasey, E., . . . Iriarte, J. (2017). Evidence for mid-Holocene rice domestication in the Americas. Nature Ecology & Evolution, 1(11), 1693-1698. doi: 10.1038/s41559-017-0322-4
» https://doi.org/10.1038/s41559-017-0322-4 - Iriarte, J., & Dickau, R. (2012). Las culturas del maíz? Arquebotânica de las sociedades hidráulicas de las tierras bajas sudamericanas. Amazônica, 4, 30-58.
- Juggins, S. (2010). C2 Software (Versão 1.6) [software de computador]. Austrália. Recuperado de https://www.staff.ncl.ac.uk/stephen.juggins/software/C2Home.htm
» https://www.staff.ncl.ac.uk/stephen.juggins/software/C2Home.htm - Kater, T. (2018). O sítio Teotônio e as reminiscências de uma longa história indígena no alto Rio Madeira (Mestrado em Arqueologia). Universidade Federal de Sergipe, Sergipe, Aracajú.
- Kater, T. (2020). A temporalidade das ocupações ceramistas no sítio Teotônio. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, 15(2), e20190078. doi: 10.1590/2178-2547-BGOELDI-2019-0078
» https://doi.org/10.1590/2178-2547-BGOELDI-2019-0078 - Kater, T., Almeida, F. Almeida, F. O, Mongeló, G. Z.; Watling, J.; Neves, E. G. (2020). Variabilidade estratigráfica e espacial dos contextos cerâmicos no sítio Teotônio. Revista de Arqueologia, 33(1), 198-220.
- Katz, O., Cabanes, D., Weiner, S., Maeir, A. M., Boaretto, E., & Shahack-Gross, R. (2010). Rapid phytolith extraction for analysis of phytolith concentrations and assemblages during an excavation: an application at Tell es-Safi/Gath, Israel. Journal of Archaeological Science, 37(7), 1557-1563. doi: 10.1016/j.jas.2010.01.016
» https://doi.org/10.1016/j.jas.2010.01.016 - Killion, T. W. (2013). Nonagricultural cultivation and social complexity: the Olmec, their ancestors, and Mexico’s Southern Gulf Lowlands. Current Anthropology, 54(5), 569-606. doi: 10.1086/673140
» https://doi.org/10.1086/673140 - Kistler, L., Maezumi, S. Y., Gregorio de Souza, J., Przelomska, N. A., Malaquias Costa, F., Smith, O., Loiselle, H., . . . Allaby, R. G. (2018). Multiproxy evidence highlights a complex evolutionary legacy of maize in South America. Science, 362(6420), 1309-1313. doi: http://doi.org/10.1126/science.aav0207
» https://doi.org/10.1126/science.aav0207 - Lathrap, D. W. (1970). The Upper Amazon London: Thames and Hudson.
- Levis, C., Costa, F. R. C., Bongers, F., Peña-Claros, M., Clement, C. R., Junqueira, A. B., . . . Ter Steege, H. (2017). Persistent effects of pre-Columbian plant domestication on Amazonian forest composition. Science, 355(6328), 925-931. doi: 10.1126/science.aal0157
» https://doi.org/10.1126/science.aal0157 - Levis, C., Flores, B. M., Moreira, P. A., Luize, B. G., Alves, R. P., Franco-Moraes, J., . . . Clement, C. R. (2018). How people domesticated Amazonian forests. Frontiers in Ecology and Evolution, 5(171). doi: 10.3389/fevo.2017.00171
» https://doi.org/10.3389/fevo.2017.00171 - Lombardo, U., Ruiz-Pérez, J., & Madella, M. (2016). Sonication improves the efficiency, efficacy and safety of phytolith extraction. Review of Palaeobotany and Palynology, 235, 1-5. doi: 10.1016/j.revpalbo.2016.09.008
» https://doi.org/10.1016/j.revpalbo.2016.09.008 - Madella, M., Alexandre, A., & Ball, T., 2005. International Code for Phytolith Nomenclature 1.0. Annals of Botany, 96(2), 253-260. doi: https://doi.org/10.1093/aob/mci172
» https://doi.org/10.1093/aob/mci172 - Maezumi, S. Y., Alves, D., Robinson, M., Gregorio de Souza, J., Levis, C., Barnett, R. L., . . . Iriarte, J. (2018). The legacy of 4,500 years of polyculture agroforestry in the eastern Amazon. Nature Plants, 4(8), 540-547. doi: 10.1038/s41477-018-0205-y
» https://doi.org/10.1038/s41477-018-0205-y - Meggers, B. J. (1954). Environmental limitations on the development of culture. American Anthropologist, 56, 801-824.
- Miller, E. T. [outros não especificados]. (1992). Arqueologia nos empreendimentos hidreléctricos da Electronorte: resultados preliminares Brasília: Electronorte.
- Mongeló, G. Z. (2015). O Formativo e os modos de produção: ocupações pré-ceramistas no alto Rio Madeira - RO (Mestrado em Arqueologia). Universidade de São Paulo, São Paulo.
- Mongeló, G. Z. (2020). Ocupações humanas do Holoceno inicial e médio no sudoeste amazônico. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, 15(2), e20190079. doi: 10.1590/2178-2547-BGOELDI-2019-0079
» https://doi.org/10.1590/2178-2547-BGOELDI-2019-0079 - Moraes, C. P. (2015). O determinismo agrícola na arqueologia amazônica. Estudos Avançandos, 29(83), 25-43. doi: http://dx.doi.org/10.1590/S0103-40142015000100004
» https://doi.org/10.1590/S0103-40142015000100004 - Morcote-Ríos, G., Bernal, R., & Raz, L. (2016). Phytoliths as a tool for archaeobotanical, palaeobotanical and palaeoecological studies in Amazonian palms. Botanical Journal of the Linnean Society, 182(2), 348-360. doi: https://doi.org/10.1111/boj.12438
» https://doi.org/10.1111/boj.12438 - Neves, E. G. (2006). Arqueologia da Amazônia Rio de Janeiro: Jorge Zahar.
- Neves, E. G. (2007). El Formativo que nunca terminó: la larga historia de estabilidad en las ocupaciones humanas de la Amazonía Central. Boletín de Arqueología PUCP, 11, 117-142.
- Neves, E. G. (2013). Was agriculture a key productive activity in pre-Colonial Amazonia? The stable productive basis for social equality in the Central Amazon. In E. S. Brondízio & E. F. Moran (Orgs.), Human-Environment Interactions: current and future decisions (pp. 371-388). Dordrecht: Springer.
- Neves, E. G. (2014). La incipiencia permanente: la Amazonia bajo el insistente destino de la incompletitud. In M. Campagno (Org.), Pierre Clastres y Las Sociedades Antiguas (1. ed., pp. 65-80). Buenos Aires: Miño y Dávila.
- Neves, E. G., Guapindaia, V. L. C., Lima, H. P., Costa, B. L. S., & Gomes, J. (2014). A tradição Pocó-Açutuba e os primeiros sinais visíveis de modificações de paisagens na calha do Amazonas. In S. Rostain (Org.), Antes de Orellana: Actas del 3er Encuentro Internacional de Arqueología Amazônica (pp. 137-158). Quito: IFEA.
- Oliver, J. R. (2008). The archaeology of agriculture in ancient Amazonia. In H. Silverman & W. H. Isbell (Orgs.), Handbook of South American Archaeology (pp. 185-216). New York: Springer.
- Olsen, K. M., & Schaal, B. A. (1999). Evidence on the origin of cassava - phylogeography of Manihot esculenta Proceedings of the National Academy of Sciences of the United States of America, 96(10), 5586-5591. doi: 10.1073/pnas.96.10.5586
» https://doi.org/10.1073/pnas.96.10.5586 - Perry, L. (2002). Starch analyses reveal multiple functions of quartz “manioc” grater flakes from the Orinoco Basin, Venezuela. Interciencia, 27(11), 635-639.
- Piperno, D. R. (2011). The origins of plant cultivation and domestication in the New World tropics: patterns, process, and new developments. Current Anthropology, 52(S4), S453–S470. doi: 10.1086/659998
» https://doi.org/10.1086/659998 - Piperno, D. R., & Dillehay, T. D. (2008). Starch grains on human teeth reveal early broad crop diet in northern Peru. Proceedings of the National Academy of Sciences, 105(50), 19622-19627. doi: 10.1073/pnas.0808752105
» https://doi.org/10.1073/pnas.0808752105 - Roosevelt, A. C. (1980). Parmana: prehistoric maize and manioc subsistence along the Amazon and Orinoco New York: Academic Press.
- Schmidt, M. J., Py-Daniel, A. R., Morães, C. P., Valle, R. B. M., Caromano, C. F., Texeira, W. G., . . . Heckenberger, M. J. (2014). Dark earths and the human built landscape in Amazonia: a widespread pattern of anthrosol formation. Journal of Archaeological Science, 42, 152-165. doi: 10.1016/j.jas.2013.11.002
» https://doi.org/10.1016/j.jas.2013.11.002 - The International Code for Starch Nomenclature (ICSN). (2011). Recuperado de http://fossilfarm.org/ICSN/Code.html
» http://fossilfarm.org/ICSN/Code.html - Torrence, R., & Barton, H. (2006). Ancient Starch Research Walnut Creek, California: Left Coast Press Inc.
- Vinton, S. D., Perry, L., Reinhard, K. J., Santoro, C. M., & Teixeira-Santos, I. (2009). Impact of empire expansion on household diet: the Inka in northern Chile’s Atacama Desert. PLoS ONE, 4(11), 1-5. doi: 10.1371/journal.pone.0008069
» https://doi.org/10.1371/journal.pone.0008069 - Watling, J., Shock, M. P., Mongeló, G. Z., Almeida, F. O., Kater, T., De Oliveira, P. E., & Neves, E. G. (2018). Direct archaeological evidence for Southwestern Amazonia as an early plant domestication and food production centre. PLos ONE, 13(7), 1-28. doi: 10.1371/journal.pone.0199868
» https://doi.org/10.1371/journal.pone.0199868 - Watling, J., Castro, M. T., Simon, M. F., Rodrigues, F. O., Brilhante de Medeiros, M., De Oliveira, P. E, Neves, E. G. (2020). Phytoliths from native plants and surface soils in the Upper Madeira basin, SW Amazonia, and their potential for paleoecological reconstruction. Quaternary International doi: https://doi.org/10.1016/j.quaint.2020.03.045.
» https://doi.org/10.1016/j.quaint.2020.03.045 - Zuse, S. (2014). Variabilidade cerâmica e diversidade cultural no alto rio Madeira, Rondônia (Doutorado em Arqueologia). Universidade de São Paulo, São Paulo.
- Zuse, S. (2016). Variabilidade cerâmica e diversidade cultural no alto rio Madeira. In Barreto, C., Lima, H. P., & Jaimes Betancourt, C. (Orgs.), Cerâmicas arqueológicas da Amazônia: rumo a uma nova síntese (pp. 385-401). Belém: IPHAN.
- Zuse, S. (2020). Tecnologias cerâmicas no alto rio Madeira: síntese, cronologia e perspectivas. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, 15(2), e20190082. doi: 10.1590/2178-2547-BGOELDI-2019-0082
» https://doi.org/10.1590/2178-2547-BGOELDI-2019-0082 - Watling, J., Almeida, F., Kater, T., Zuse, S., Shock, M. P., Mongeló, G., Bespalez, E., Santi, J. R., & Neves, E. G. (2020). Arqueobotânica das ocupações ceramistas na Cachoeira do Teotônio. Boletim do Museu Paraense Emílio Goeldi. Ciências Humanas, 15(2), e20190075. doi: 10.1590/2178-2547-BGOELDI-2019-0075
» https://doi.org/10.1590/2178-2547-BGOELDI-2019-0075
Datas de Publicação
-
Publicação nesta coleção
17 Jul 2020 -
Data do Fascículo
2020
Histórico
-
Recebido
02 Ago 2019 -
Aceito
04 Out 2019

 Desenho:
Desenho: 
 Legenda: X = presença de taxa com baixas frequências (< 2%).
Legenda: X = presença de taxa com baixas frequências (< 2%).
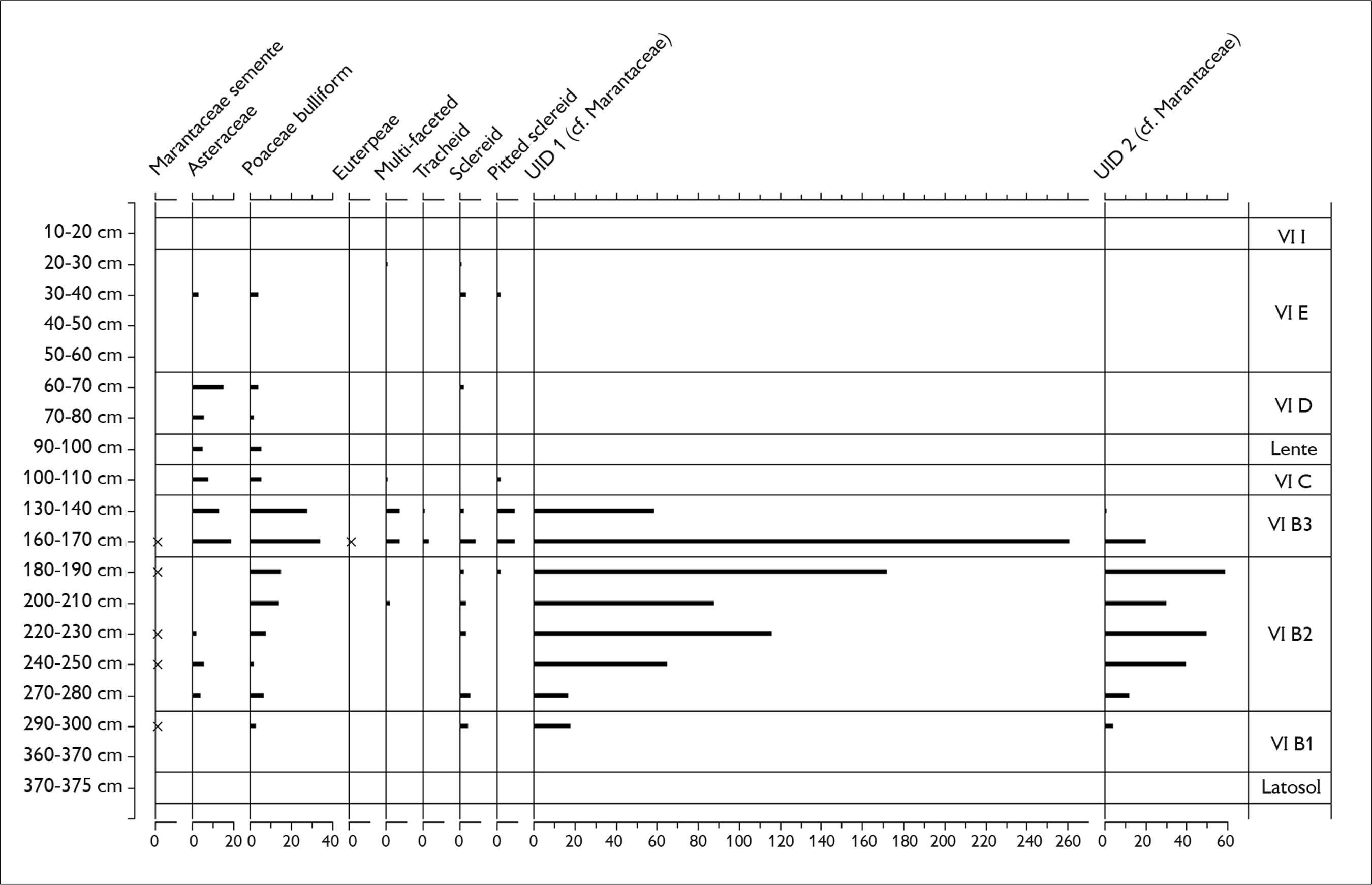 Legenda: X = presença de taxa com baixas frequências (< 2%).
Legenda: X = presença de taxa com baixas frequências (< 2%).
 Legenda: X = presença de taxa com baixas frequências (< 2%).
Legenda: X = presença de taxa com baixas frequências (< 2%).
 Legenda: X = presença de taxa com baixas frequências (< 2%).
Legenda: X = presença de taxa com baixas frequências (< 2%).