
Abstract
The effect of water extracts of doenjang (WED) on cyclooxygenase pathway and cytokine production in lipopolysaccharide (LPS)-treated RAW 264.7 macrophage cells was investigated. WED was fractionated by ultrafiltration into five individual groups of molecular weights. All of the WED fractions significantly inhibited the production of prostaglandin E2 (PGE2) and expression of cyclooxygenase-2 (COX-2) in LPS-stimulated RAW 264.7 macrophages. Furthermore, each of the WED fractions significantly decreased production of pro-inflammatory cytokine such as tumor necrosis factor-α (TNF-α) and interleukin-1β (IL-1β) (p<0.05). The fraction between 10 to 30 kDa of WED showed the highest inhibition of pro-inflammatory cytokine production, LPS-induced COX-2 expression and PEG2 production. These results suggest that WED fractions may have the anti-inflammatory effect via the stimulus of COX-2 and TNF-α in LPS-stimulated inflammation in RAW 264.7 cells.
Keywords:
water extracts of doenjang (WED); fermented soybean paste; oxidative stress; inflammation; Lipopolysaccharide
1 Introduction
Inflammation plays a central role in various chronic diseases, such as cardiovascular disease, cancer, asthma, rheumatoid arthritis and bronchitis (Heo et al., 2010Heo, S. J., Yoon, W. J., Kim, K. N., Ahn, G. N., Kang, S. M., Kang, D. H., Affan, A., Oh, C., Jung, W. K., & Jeon, Y. J. (2010). Evaluation of anti-inflammatory effect of fucoxanthin isolated from brown algae in lipopolysaccharide-stimulated RAW 264.7 macrophages. Food and Chemical Toxicology, 48(8-9), 2045-2051. http://dx.doi.org/10.1016/j.fct.2010.05.003. PMid:20457205.
http://dx.doi.org/10.1016/j.fct.2010.05....
; Lee et al., 2012bLee, S. J., Kim, E. K., Kim, Y. S., Hwang, J. W., Lee, K. H., Choi, D. K., Kang, H., Moon, S. H., Jeon, B. T., & Park, P. J. (2012b). Purification and characterization of a nitric oxide inhibitory peptide from Ruditapes philippinarum. Food and Chemical Toxicology, 50(5), 1660-1666. http://dx.doi.org/10.1016/j.fct.2012.02.021. PMid:22386812.
http://dx.doi.org/10.1016/j.fct.2012.02....
). Among inflammatory processes, macrophages act a major role in inflammation-related diseases and secrete pro-inflammatory cytokines such asinterleukin-1 beta (IL-1β), IL-6 and tumor necrosis factor-alpha (TNF-α) and inflammation mediators including nitric oxide (NO) and prostaglandins (Heo et al., 2010Heo, S. J., Yoon, W. J., Kim, K. N., Ahn, G. N., Kang, S. M., Kang, D. H., Affan, A., Oh, C., Jung, W. K., & Jeon, Y. J. (2010). Evaluation of anti-inflammatory effect of fucoxanthin isolated from brown algae in lipopolysaccharide-stimulated RAW 264.7 macrophages. Food and Chemical Toxicology, 48(8-9), 2045-2051. http://dx.doi.org/10.1016/j.fct.2010.05.003. PMid:20457205.
http://dx.doi.org/10.1016/j.fct.2010.05....
). Moreover, inflammation-related enzymes such as inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) are associated with the growth of numerous inflammatory diseases (Murakami & Ohigashi, 2007Murakami, A., & Ohigashi, H. (2007). Targeting NOX, INOS and COX-2 in inflammatory cells: chemoprevention using food phytochemicals. International Journal of Cancer, 121(11), 2357-2363. http://dx.doi.org/10.1002/ijc.23161. PMid:17893865.
http://dx.doi.org/10.1002/ijc.23161...
). Therefore, inhibition of inflammaproduction of inflammatory mediator is the important therapeutic target for healing of inflammation-related diseases (Ko & Jeon, 2015Ko, S. C., & Jeon, Y. J. (2015). Anti-inflammatory effect of enzymatic hydrolysates from Styela clava flesh tissue in lipopolysaccharide-stimulated RAW 264.7 macrophages and in vivo zebrafish model. Nutrition Research and Practice, 9(3), 219-226. http://dx.doi.org/10.4162/nrp.2015.9.3.219. PMid:26060532.
http://dx.doi.org/10.4162/nrp.2015.9.3.2...
).
Food proteins from natural sources have been used to obtain a various range of bioactive peptides (Hou et al., 2014Hou, Y., Liu, W., Cheng, Y., Zhou, J., Wu, L., & Yang, G. (2014). Production optimization and characterization of immunomodulatory peptides obtained from fermented goat placenta. Food Science and Technology, 34(4), 723-729. http://dx.doi.org/10.1590/1678-457X.6448.
http://dx.doi.org/10.1590/1678-457X.6448...
; Yoshikawa et al., 2000Yoshikawa, M., Fujita, H., Matoba, N., Takenaka, Y., Yamamoto, T., Yamauchi, R., Tsuruki, H., & Takahata, K. (2000). Bioactive peptides derived from food proteins preventing lifestyle-related diseases. BioFactors, 12(1-4), 143-146. http://dx.doi.org/10.1002/biof.5520120122. PMid:11216476.
http://dx.doi.org/10.1002/biof.552012012...
). Bioactive peptides usually contain short peptides (2-20 amino acid) derived from protein, but in some cases, they may consist of more than 20 amino acids (Yoshikawa et al., 2000Yoshikawa, M., Fujita, H., Matoba, N., Takenaka, Y., Yamamoto, T., Yamauchi, R., Tsuruki, H., & Takahata, K. (2000). Bioactive peptides derived from food proteins preventing lifestyle-related diseases. BioFactors, 12(1-4), 143-146. http://dx.doi.org/10.1002/biof.5520120122. PMid:11216476.
http://dx.doi.org/10.1002/biof.552012012...
; Kitts & Weiler, 2003Kitts, D. D., & Weiler, K. (2003). Bioactive proteins and peptides from food sources. Applications of bioprocesses used in isolation and recovery. Current Pharmaceutical Design, 9(16), 1309-1323. http://dx.doi.org/10.2174/1381612033454883. PMid:12769739.
http://dx.doi.org/10.2174/13816120334548...
). They are inert within the sequence of their parent protein and may be released by enzymatic proteolysis such as during gastrointestinal digestion or during food processing. Many of the well-known bioactive peptides are multifunctional and can apply more than one of the physiological effects mentioned (Korhonen & Pihlanto, 2003Korhonen, H., & Pihlanto, A. (2003). Food-derived bioactive peptides-opportunities for designing future foods. Current Pharmaceutical Design, 9(16), 1297-1308. http://dx.doi.org/10.2174/1381612033454892. PMid:12769738.
http://dx.doi.org/10.2174/13816120334548...
).
Soybean (glycine max) is a source of a various of bioactive substances, including dietary fiber, oligosaccharides, isoflavones, phytic acid, saponin, protein and protein hydrolysates, unsaturated fatty acids, and phenol compounds (Choi & Rhee, 2006Choi, M. S., & Rhee, K. C. (2006). Production and Processing of soybeans and nutrition and safety of isoflavone and other soy products for human health. Journal of Medicinal Food, 9(1), 1-10. http://dx.doi.org/10.1089/jmf.2006.9.1. PMid:16579721.
http://dx.doi.org/10.1089/jmf.2006.9.1...
). Soybean protein and its hydrolysates (peptides) exhibit various biological activities such as suppression of anticancer (de Lumen, 2005Lumen, B. O. (2005). Lunasin: A cancer preventive peptide. Nutrition Reviews, 63(1), 16-21. http://dx.doi.org/10.1111/j.1753-4887.2005.tb00106.x. PMid:15730231.
http://dx.doi.org/10.1111/j.1753-4887.20...
) and antihypertensive (Mallikarjun Gouda et al., 2006Mallikarjun Gouda, K. G., Gowda, L. R., Rao, A. G., & Prakash, V. (2006). Angiotensin I-converting enzyme inhibitory peptide derived from glycinin, the 11S globulin of soybean (Glycine max). Journal of Agricultural and Food Chemistry, 54(13), 4568-4573. http://dx.doi.org/10.1021/jf060264q. PMid:16786999.
http://dx.doi.org/10.1021/jf060264q...
) properties and immunostimulation (Tsuruki et al., 2003Tsuruki, T., Kishi, K., Takahashi, M., Tanaka, M., Matsukawa, T., & Yoshikawa, M. (2003). Soymetide, an immunostimulating peptide derived from soybean β-conglycinin, is an fMLP agonist. FEBS Letters, 540(1-3), 206-210. http://dx.doi.org/10.1016/S0014-5793(03)00265-5. PMid:12681509.
http://dx.doi.org/10.1016/S0014-5793(03)...
).
Specifically, soybean hydrolysates have produced a number of bioactive peptides showing anti-inflammatory effects on macrophage cell lines with preparations from germinated beans eliciting the higher responses (Vernaza et al., 2012Vernaza, M. G., Dia, V. P., de Mejia, E. G., & Chang, Y. K. (2012). Antioxidant and antiinflammatory properties of germinated and hydrolysed Brazilian soybean flours. Food Chemistry, 134(4), 2217-2225. http://dx.doi.org/10.1016/j.foodchem.2012.04.037. PMid:23442677.
http://dx.doi.org/10.1016/j.foodchem.201...
). The soybean-derived peptide lunasin consists of a unique 43 amino acids (approximately 4.8 kD) and refers to exert anti-inflammatory effects including COX-2 levels NF-𝜅B activity, reduced cytokine expression and suppression of NF-𝜅B activity (Hernández-Ledesma et al., 2009Hernández-Ledesma, B., Hsieh, C. C., & de Lumen, B. O. (2009). Antioxidant and anti-inflammatory properties of cancer preventive peptide lunasin in RAW 264.7 macrophages. Biochemical and Biophysical Research Communications, 390(3), 803-808. http://dx.doi.org/10.1016/j.bbrc.2009.10.053. PMid:19836349.
http://dx.doi.org/10.1016/j.bbrc.2009.10...
). The presence of an RGD (Arg-Gly-Asp) motif in lunasin and similar peptides is believed to contribute to their anti-inflammatory effects, involving antagonism of downstream pro-inflammatory cascades and integrin signaling (Cam & Mejia, 2012Cam, A., & Mejia, E. G. (2012). RGD-peptide lunasin inhibits Akt-mediated NF-κB activation in human macrophages through interaction with the αVβ3 integrin. Molecular Nutrition & Food Research, 56(10), 1569-1581. http://dx.doi.org/10.1002/mnfr.201200301. PMid:22945510.
http://dx.doi.org/10.1002/mnfr.201200301...
). Gonzalez de Mejia & Dia (2009)Mejia, E. G., & Dia, V. P. (2009). Lunasin and lunasin-like peptides inhibit inflammation through suppression of NF-κB pathway in the macrophage. Peptides, 30(12), 2388-2398. http://dx.doi.org/10.1016/j.peptides.2009.08.005. PMid:19682518.
http://dx.doi.org/10.1016/j.peptides.200...
demonstrated that the mixture of three peptides with molecular weight of 5, 8, and 14 kDa isolated from defatted soybean flour inhibited inflammatory response by suppressing the iNOS/NO and COX-2/PGE2 pathways.
Soybean-based fermented foods such as whole cooked soybean fermented product (chungkookjang) and soybean paste (doenjang) are very popular in Korea (Lee, 2012Lee, S. H. (2012). Physiological activities of water extracts of doenjang fractionated to their molecular weight of nitrogenous compounds (Ph.D. Dissertation). Inje University, Gimhae, Gyeongnam, Korea.). Chungkookjang has been made by fermenting with microorganism with completely cooked soybean without salt addition. To make doenjang, soybean is cooked, crushed and molded to obtain meju (the shape of a brick) at the beginning of winter. The meju is dried and fermented in the open air. Thus, meju performs as a medium for solid culture. The fermented meju is then aged in salted water for a period of longer than 6 months. The non-aqueous slurry separated from the aged solution is used as doenjang (Lee, 2012Lee, S. H. (2012). Physiological activities of water extracts of doenjang fractionated to their molecular weight of nitrogenous compounds (Ph.D. Dissertation). Inje University, Gimhae, Gyeongnam, Korea.). Therefore, various hydrolysates of biological substances, including peptides, are present in doenjang (Lee et al., 2014Lee, S. H., Jung, J. H., Lee, H. A., Lee, H.-S., Jeong, H. D., & Cherl, J. (2014). Fractionation of nitrogenous compounds from water extracts of doenjang and their physiological activities. Food Science and Biotechnology, 23(2), 523-529. http://dx.doi.org/10.1007/s10068-014-0072-x.
http://dx.doi.org/10.1007/s10068-014-007...
).
Few studies have been carried out the anti-inflammatory activity of peptides derived from Korean fermented soybean products (Lee et al., 2014Lee, S. H., Jung, J. H., Lee, H. A., Lee, H.-S., Jeong, H. D., & Cherl, J. (2014). Fractionation of nitrogenous compounds from water extracts of doenjang and their physiological activities. Food Science and Biotechnology, 23(2), 523-529. http://dx.doi.org/10.1007/s10068-014-0072-x.
http://dx.doi.org/10.1007/s10068-014-007...
). Hwang et al. (2011)Hwang, J. S., Yoo, H. J., Song, H. J., Kim, K. K., Chun, Y. J., Matsui, T., & Kim, H. B. (2011). Inflammation related signaling pathways implicating TGF𝛽 are revealed in the expression profiling of MCF7 cell treated with fermented soybean, Chungkookjang. Nutrition and Cancer, 63(4), 645-652. http://dx.doi.org/10.1080/01635581.2011.551987. PMid:21547849.
http://dx.doi.org/10.1080/01635581.2011....
demonstrated that a bioactive peptide from chungkookjang exhibited anti-inflammatory effects in breast cancer cells by activation of transforming growth factor (TGF)-beta signaling and down-regulation of cytokine/chemokine expression. An inhibitory effect on the NO production in LPS-stimulated cells was observed in fractions of nitrogenous compounds separated by molecular weight level from water extracts of doenjang (WED) (Lee et al., 2014Lee, S. H., Jung, J. H., Lee, H. A., Lee, H.-S., Jeong, H. D., & Cherl, J. (2014). Fractionation of nitrogenous compounds from water extracts of doenjang and their physiological activities. Food Science and Biotechnology, 23(2), 523-529. http://dx.doi.org/10.1007/s10068-014-0072-x.
http://dx.doi.org/10.1007/s10068-014-007...
). It is not still clear yet whether the anti-inflammatory effect of WED fractions is associated with the inflammatory mediators or not. Therefore, the aim of study is to investigate the effect of WED fractions on ROS production and expression of pro-inflammatory cytokines in LPS-stimulated RAW 264.7 macrophage cells.
2 Materials and methods
2.1 Materials
Traditional doenjang was obtained from the Korean local areas of Oksan-seowon, Gyeongju, Gyeongsangbuk-do (Lee et al., 2014Lee, S. H., Jung, J. H., Lee, H. A., Lee, H.-S., Jeong, H. D., & Cherl, J. (2014). Fractionation of nitrogenous compounds from water extracts of doenjang and their physiological activities. Food Science and Biotechnology, 23(2), 523-529. http://dx.doi.org/10.1007/s10068-014-0072-x.
http://dx.doi.org/10.1007/s10068-014-007...
). The doenjang was freeze dried and stored at 4 °C. 6-carboxy-2', 7'-dichlorofluorescein diacetate (DCFH-DA) was provided from Molecular Probes (Eugene, OR, USA). Lipopolysaccharides (LPS), phosphoric acid, sodium nitrite, naphthylethylenediamine dihydrochloride (NED) and sulfanilamide were purchased from Sigma Chemical Co. (St. Louis, MO, USA). A 3-(4,5-dimethylthiazol-2-yl]-5-(3-carboxy-methoxyphenyl)-2-(4-sulfenyl)-2H-tetrazolium (MTS) assay kit was obtained from Promega (Madison, WI, USA). The goat anti-rabbit IgG-HRP and goat anti-mouse IgG-HRP were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). The protein extraction solution was provided from Intron Biotechnology (Gyeonggi-do, Korea). The PGE2 ELISA kit was provided from R&D Systems (Minneapolis, MI, USA). The polyvinylidene fluoride membrane and ECL kit were provided from Amersham Pharmacia Biotech (Buckinghamshire, UK). TNF-α and IL-1β ELISA kits were obtained from Pierce Biotechnology (Rockford, IL, USA). The anti-iNOS, anti-COX-2 and anti-β-actin antibodies were provided from Cell Signaling Technology (Beverly, MA, USA). All other chemicals and reagents were of analytical grade.
2.2 Sample preparation
WED and WED fractions according to their molecular weight were prepared according to a previously described method (Lee et al., 2014Lee, S. H., Jung, J. H., Lee, H. A., Lee, H.-S., Jeong, H. D., & Cherl, J. (2014). Fractionation of nitrogenous compounds from water extracts of doenjang and their physiological activities. Food Science and Biotechnology, 23(2), 523-529. http://dx.doi.org/10.1007/s10068-014-0072-x.
http://dx.doi.org/10.1007/s10068-014-007...
). The freeze-dried doenjang (100 g) was mixed with 10 volumes (w/v) of distilled water for 1 h at room temperature, and water-soluble constituents were isolated using filter paper (No. 40, Whatman, Maidstone, UK). The water-soluble constituents were desalted (WED) by FPLC gel filtration with a Fast Desalting column (Pharmacia, USA), freeze-dried, and stored at -20 °C. Ten grams of freeze-dried WED was liquefied in 5 volumes of distilled water and separated into 5 groups WED fractions with molecular weights less than 1 (WEDI), between 1 and 3 (WEDII), 3 and 5 (WEDIII), 5 and 10 (WEDIV), and 10 and 30 kDa (WEDV) using ultrafiltration membranes (Amicon, Lexington, USA). All of the separated fractions were freeze-dried and kept under -20 °C until used.
2.3 Cell culture
RAW264.7 cells and mouse macrophage cell line were purchased from KCLB (KCLB; Seoul, Korea). Cells were cultured in DMEM (GIBCO, Grand Island, NY, USA) prepared with 10% fetal bovine serum (FBS) and 1% penicillin-streptomycin. Cells from passages 20 to 25 were used and maintained in an incubator at 37 °C in 5% CO2. RAW 264.7 cells were cultivated to 80-90% confluence and performed quiescent by starvation for at least 24 h.
2.4. Cell viability
A Cell Titer 96 colorimetric assay (Promega, Madison, WI, USA) using MTS reagent determined cell viability. RAW 264.7 cells (1x105) were distributed into wells of a 96-well plate and added with WED and WED fractions for 1 h before 100 ng/mL LPS treatment. After 19 h of cultivation, 20 μL MTS reagent added. Cells were cultivated for another 4 h at 37 oC in 5% CO2. An absorbance at 490 nm was measured using a microplate reader (Molecular Devices, Sunnyvale, CA, USA).
2.5 Reactive Oxygen Species (ROS) Generation in LPS-stimulated RAW 264.7 Cells
Intracellular ROS generations were determined using the DCFH-DA staining method (Kalyanaraman et al., 2012Kalyanaraman, B., Darley-Usmar, V., Davies, K. J., Dennery, P. A., Forman, H. J., Grisham, M. B., Mann, G. E., Moore, K., Roberts, L. J. 2nd, & Ischiropoulos, H. (2012). Measuring reactive oxygen and nitrogen species with fluorescent probes: challenges and limitations. Free radical biology and medicine, 52(1), 1-6. http://dx.doi.org/10.1016/j.freeradbiomed.2011.09.030.
http://dx.doi.org/10.1016/j.freeradbiome...
). Briefly, RAW 264.7 cells (1x105) were distributed into wells of a 96-well plate and supplemented with WED and WED fractions for 1 h before 1μg/mL LPS treatment and then incubated for 24 h. After 30 min of treatment, 10 μM DCFH-DA was added and cultivated in the dark. Incubated cells were washed three times using phosphate-buffered saline (PBS). ROS-mediated fluorescence with excitation and emission wavelengths of 485 nm and 528 nm was measured using a fluorescence microplate reader (Molecular Device, Sunnyvale, CA, USA).
2.6 NO Production in LPS-stimulated RAW 264.7 Cells
RAW 264.7 cells (1x105) were pretreated with WED and WED fractions for 1 h before being incubated with 1μg/mL LPS for 24 h. The nitrite concentration in the culture medium was measured as an amount of NO production using Griess reagent system. Fifty μL of each supernatant was added and mixed with 50 μL of sulfanilamide solution and then cultivated for 10 min at room temperature protected from light. The same volume of NED (N-1-napthylethylenediamine dihydrochloride) solution was added and cultivated for 10 min at room temperature in dark. Absorbance of the mixture solution at 540 nm was measured using microplate plate reader (Molecular Devices, Sunnyvale, CA, USA)
2.7 Enzyme-linked Immunosorbent Assay (ELISA) Analysis
RAW 264.7 cells (1x105) were distributed into wells of a 24-well plate and prepared with WED and WED fractions and stabilized for 1 h, after which they were induced with 1 μg/mL LPS. The supernatants were subsequently separated and the amount of PGE2, TNF-α and IL-1β production was measured using an ELISA kit.
2.8 Immunoblot analysis
RAW 264.7 cells (1x105) were pre-incubated with WED and WED fractions for 1 h before being stimulated with 1 μg/mL LPS. After 23 h of incubation, the cells were washed twice using ice-cold PBS and gathered by scraping in 1 mL of ice-cold PBS. Whole cell lysates were extracted by Pro-prep protein extraction solution (Intron Biotechnology, Gyeonggi-do, Korea). Protein concentration was analyzed with a commercial protein assay kit (Bio-Rad, Hercules, CA, USA). Equal protein concentrations were subjected to 10% SDS-polyacrylamide gel electrophoresis and transferred onto a polyvinylidene fluoride membrane (GE Healthcare, Piscataway, NJ). After blocking with PBS containing 5% non-fat dry milk for 1 h at room temperature, each membrane was incubated with a specific primary antibody overnight at 4ºC, including anti-cyclooxygenase-2 (Cox2, 1:1,000), anti-iNOS (CD31, 1:1,000), anti-tumor necrosis factor-α (TNFα, 1:1,000) and anti-interleukin 1β (IL-1 β, 1:1,000, all from Santa Cruz Biotechnology, Santa Cruz, CA). Each membrane was washed twice with PBS containing 0.1% Tween-20 (PBST), immunoblotted with horseradish peroxidase-conjugated anti-mouse or anti-rabbit for 1 h, washed three times in PBST, and visualized by enhanced ECL (Amersham Pharmacia Biotech, Buckinghamshire, UK). The band intensities was determine using the Fusion Fx image acquisition system (Vilber Lourmat, Torcy, France).
2.9 Statistical analysis
The data were determined with SPSS version 22.0 (SPSS, Chicago, IL, USA) and are indicated as the means ± standard deviations (SDs). The differences between the groups were analyzed using 1-way ANOVA, and statistical significance was defined at p < 0.05 by Tukey’s test.
3 Results and discussion
3.1 Proximate composition and distribution of nitrogenous compounds in WED and WED fractions
Proximate contents (wt. % in dry basis) of protein, carbohydrate and ash in WED were 78.0, 15.7 and 6.3%, respectively. Most of the nitrogenous compounds were amino acids and peptide. Distribution ratio of nitrogenous compounds in five fractions separated by their molecular weight range were 92.2 (WEDI), 3.0 (WEDII), 2.1 (WEDIII), 0.0 (WDEIV) and 2.6% (WEDV). A fraction of 5 to 10 kDa molecular weight (WEDIV) has a very small proportion of nitrogenous compounds. The free amino acid concentration was 3410.9 ± 32.9 mg%, which consists of 59.8% of the total amino acids in WEDI (Lee, 2012Lee, S. H. (2012). Physiological activities of water extracts of doenjang fractionated to their molecular weight of nitrogenous compounds (Ph.D. Dissertation). Inje University, Gimhae, Gyeongnam, Korea.; Lee et al., 2014Lee, S. H., Jung, J. H., Lee, H. A., Lee, H.-S., Jeong, H. D., & Cherl, J. (2014). Fractionation of nitrogenous compounds from water extracts of doenjang and their physiological activities. Food Science and Biotechnology, 23(2), 523-529. http://dx.doi.org/10.1007/s10068-014-0072-x.
http://dx.doi.org/10.1007/s10068-014-007...
).
3.2 Effect of the WED and WED fractions on ROS generation in LPS-stimulated RAW 264.7 cells
Each fraction of WED was applied to the LPS-stimulated RAW 264.7 cell in order to examine cytotoxicity of the WED. The dose of the critical concentration revealing cytotoxicity on the RAW 264.7 cell with and without treatment of LPS was 400 μg/mL for WED and WED fractions, except for molecular weights more than 10 kDa. A fraction with molecular weight more than 10 kDa showed cytotoxicity at a concentration of 100 μg/mL (Figure 1). No significant differences in the effect of WED and WED fractions on cell viabilities of both the RAW 264.7 and LPS-stimulated RAW 264.7 cells were found.

Effect of the WED and WED fractions on cell viability in RAW 264.7 cells. The cell viability was measured by MTS assay. RAW 264.7 cells were pretreated with different concentrations of WED fraction for 1 hr before being cultivated with or without 1 μg/mL LPS for 24 hrs. Values are mean±S.D. (n=3).
Treatment of LPS on the RAW 264.7 cell induced 3.2 times ROS generation compared to that of RAW 264.7 cell (control). The generation of ROS was inhibited by WED fractions. Particularly, a dose of 400 μg/mL on WEDII and a dose of 100 μg/mL on WEDV were significantly effective in inhibiting ROS generation compared to control (Figure 2).

Effect of the WED and WED fractions on LPS-stimulated ROS generation in RAW 264.7 cells. RAW 264.7 cells were prepared with WED and WED fractions for 1 h before being cultivated with 1 μg/mL LPS for 24 h. 10 μM DCF-DA was added during the last 30 min of the treatment. ROS generation is mean±S.D. (n=3) and expressed as fold of generation compared to control. *a-dBars with different letters differ significantly at p<0.05 by Tukey test.
ROS can influence the expression of several of genes in monocytes. H2O2 is known to activate NF-κB, which regulates the expression of pro-inflammatory cytokines and immune mediators (Lee et al., 2012aLee, H. S., Ryu, D. S., Lee, G. S., & Lee, D. S. (2012a). Anti-inflammatory effects of dichloromethane fraction from Orostachys japonicus in RAW 264.7 cells: suppression of NF-κB activation and MAPK signaling. Journal of Ethnopharmacology, 140(2), 271-276. http://dx.doi.org/10.1016/j.jep.2012.01.016. PMid:22285522.
http://dx.doi.org/10.1016/j.jep.2012.01....
). Oxidative stress is strongly associated with small cellular molecules, cellular damage, proteins, lipids, and oxidative modification of DNA by ROS. It acts a major role in a variety of common diseases and age-related degenerative conditions (Bowie & O’Neill, 2000Bowie, A., & O’Neill, L. A. (2000). Oxidative stress and nuclear factor-κB activation: a reassessment of the evidence in the light of recent discoveries. Biochemical Pharmacology, 59(1), 13-23. http://dx.doi.org/10.1016/S0006-2952(99)00296-8. PMid:10605930.
http://dx.doi.org/10.1016/S0006-2952(99)...
). Thus, dietary antioxidants capable of protecting vital molecules against oxidative damage may help prevent the onset and progression of disease (Lee et al., 2006Lee, S. O., Kim, S. Y., Han, S. M., Kim, H. M., Ham, S. S., & Kang, I. J. (2006). Corni fructus scavengers hydroxyl radicals and decreases oxidative stress in Endothelial cells. Journal of Medicinal Food, 9(4), 594-598. http://dx.doi.org/10.1089/jmf.2006.9.594. PMid:17201652.
http://dx.doi.org/10.1089/jmf.2006.9.594...
). A number of studies have been devoted to evaluating the antioxidant ability of soy protein fractions as well as to the separation and structural property of the most active peptides (Borek, 1991Borek, C. (1991). Free radical processes in multistage carcinogenesis. Free Radical Research Communications, 13(1), 745-750. http://dx.doi.org/10.3109/10715769109145854. PMid:2060846.
http://dx.doi.org/10.3109/10715769109145...
). These antioxidant peptides isolated from soybean protein hydrolysates have been identified to contain between 5 and 16 amino acids (Gibbs et al., 2004Gibbs, B. F., Zougman, A., Masse, R., & Mulligan, C. (2004). Production and characterization of bioactive peptides from soy hydrolysate and soy fermented food. Food Research International, 37(2), 123-131. http://dx.doi.org/10.1016/j.foodres.2003.09.010.
http://dx.doi.org/10.1016/j.foodres.2003...
). In this study, we demonstrated that WEDII and WEDV fractions significantly inhibited oxidative stress in LPS-activated macrophages by inhibiting ROS generation. The results suggested that WEDII with approximately 9 amino acids and WEDV with approximately 86 amino acids might contribute to the antioxidant properties. The enhancement of the antioxidant property of the fermented soybean depends on the organism used for the fermentation. It has also been shown that mixture of proteolytic microorganisms (Bacillus and fungus) during fermentation of soybean increased the total antioxidant activities (Wongputtisin et al., 2007Wongputtisin, P., Khanongnuch, C., Pongpiachan, P., & Lumyong, S. (2007). Antioxidant activity improvement of soybean meal by microbial fermentation. Research Journal of Microbiology, 2(7), 577-583. http://dx.doi.org/10.3923/jm.2007.577.583.
http://dx.doi.org/10.3923/jm.2007.577.58...
).
3.3 Effect of the WED and WED fractions on iNOS expression and NO production in LPS-stimulated RAW 264.7 cells
The effect of WED and WED fractions on LPS-stimulated NO production in RAW 264.7 cells was examined by measuring amount of nitrite released into the culture medium using the Griess reaction (Figure 3). No significant differences in NO production were found in RAW 264.7 cells treated with WED and WED fractions (Figrue 1). However, WEDII and WEDV significantly decreased LPS-stimulated NO production: 17.5 μM at 400 μg/mL and 20.58 μM at 100 μg/mL, respectively (Figure 3A).
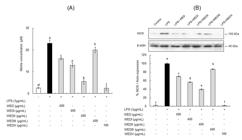
Effect of the WED and WED fractions on NO production (A) and iNOS expression (B) in RAW 264.7 cells. (A) RAW 264.7 cells were prepared with WED and WED fractions for 1 h before being cultivated with 1 μg/mL LPS for 24 h. The supernatants were isolated and measured for nitrite concentrations. (B) RAW 264.7 cells were prepared with WED and WED fractions for 1 h before being cultivated with 1 μg/mL LPS for 24 h and subjected to immunoblotting using antibody specific for iNOS. Values are mean±S.D. (n=3). *a-fBars with different letters differ significantly different at p<0.05 by Tukey test.
The results demonstrate that the NO inhibition effects of the WED and WED fractions could be brought about through the influence of iNOS protein expression. The iNOS protein expression in LPS-stimulated RAW 264.7 cells at 6 and 24 h was observed by Western blot assay. WEDII and WEDV inhibited LPS-stimulated iNOS expression by 60.4% at 400 μg/mL and 98.3% at 100 μg/mL, respectively, compared to cells treated with LPS alone (Figure 3B). The results suggest that WEDII and WEDV may contribute to preventing inflammation through inhibition of iNOS/NO pathways.
NO is a central inflammatory mediator synthesized from the amino acid by nitric oxide synthesis (NOS) (Heo et al., 2010Heo, S. J., Yoon, W. J., Kim, K. N., Ahn, G. N., Kang, S. M., Kang, D. H., Affan, A., Oh, C., Jung, W. K., & Jeon, Y. J. (2010). Evaluation of anti-inflammatory effect of fucoxanthin isolated from brown algae in lipopolysaccharide-stimulated RAW 264.7 macrophages. Food and Chemical Toxicology, 48(8-9), 2045-2051. http://dx.doi.org/10.1016/j.fct.2010.05.003. PMid:20457205.
http://dx.doi.org/10.1016/j.fct.2010.05....
). Under pathological state, NO is produced by the iNOS, which subsequently induces the tissue damage (Ko & Jeon, 2015Ko, S. C., & Jeon, Y. J. (2015). Anti-inflammatory effect of enzymatic hydrolysates from Styela clava flesh tissue in lipopolysaccharide-stimulated RAW 264.7 macrophages and in vivo zebrafish model. Nutrition Research and Practice, 9(3), 219-226. http://dx.doi.org/10.4162/nrp.2015.9.3.219. PMid:26060532.
http://dx.doi.org/10.4162/nrp.2015.9.3.2...
). Therefore, NO inhibitors are indispensable to prevention of inflammation-mediated responses. Recent studies have investigated the effect of soybean-derived hydrolysates on LPS-stimulated macrophages for the treatment of inflammation (Hernández-Ledesma et al., 2009Hernández-Ledesma, B., Hsieh, C. C., & de Lumen, B. O. (2009). Antioxidant and anti-inflammatory properties of cancer preventive peptide lunasin in RAW 264.7 macrophages. Biochemical and Biophysical Research Communications, 390(3), 803-808. http://dx.doi.org/10.1016/j.bbrc.2009.10.053. PMid:19836349.
http://dx.doi.org/10.1016/j.bbrc.2009.10...
; Cam & Mejia, 2012Cam, A., & Mejia, E. G. (2012). RGD-peptide lunasin inhibits Akt-mediated NF-κB activation in human macrophages through interaction with the αVβ3 integrin. Molecular Nutrition & Food Research, 56(10), 1569-1581. http://dx.doi.org/10.1002/mnfr.201200301. PMid:22945510.
http://dx.doi.org/10.1002/mnfr.201200301...
; Mejia & Dia, 2009Mejia, E. G., & Dia, V. P. (2009). Lunasin and lunasin-like peptides inhibit inflammation through suppression of NF-κB pathway in the macrophage. Peptides, 30(12), 2388-2398. http://dx.doi.org/10.1016/j.peptides.2009.08.005. PMid:19682518.
http://dx.doi.org/10.1016/j.peptides.200...
). NO inhibitory activity was found in every of the fractions. Therefore, many NO inhibitory substances with various molecular weight ranges were existed in the hydrolysate. Among the fractions, the most potent NO inhibition was observed in between the 10 and 30 kDa fractions (WEDV).
3.4 Effect of the WED and WED fractions on PGE2 production and COX-2 expression in LPS-stimulated RAW 264.7 cells
The effects of water extracts of doenjang (WED) on LPS-stimulated PGE2 production and COX-2 gene expression were investigated (Figure 4). Unstimulated RAW 264.7 macrophages in culture medium for 24 h produced basal amount of PGE2 (285.4 pg/mL) in the medium. After stimulation with LPS (1 μg/mL) for 24 h, the amount of PGE2 apparently increased to 1,223.2 pg/mL in medium. Addition of both WEDs and LPS to the cell strongly suppressed LPS-stimulated PGE2 production (Figure 4A). Especially, WEDV showed the strongest inhibitory activity on LPS-stimulated PGE2 production. Western blot was performed to demonstrate the effect of WED and WED fractions on COX-2 gene expression. RAW 264.7 macrophages expressed only a little amount of COX-2 protein in unstimulated cells, and WED treatment alone showed no alternation on basal COX-2 expression. Treatment of LPS (1 μg/mL) dramatically increased COX-2 protein in cells. Cotreatment of cells with LPS (1 μg/mL) and WEDV (100 μg/mL) inhibited induction of COX-2 protein in RAW 264.7 cells (Figure 4B).

The effect of the WED and WED fractions on LPS-induced PGE2 production (A) and COX-2 expression (B) in RAW 264.7 cells. (A) RAW 264.7 cells were pretreated with WED and WED fractions for 1 h before being cultivated with 1 μg/mL LPS for 24 h. The supernatants were subsequently separated, and the amount of PGE2 production was measured using an ELISA kit. (B) RAW 264.7 cells were prepared with different concentrations of WED and WED fractions for 1 h before being cultivated with 1 μg/mL LPS.
COX-2 is a major mediator of inflammation such as PGE2 and NO generation (Ahmad et al., 2002Ahmad, N., Chen, L. C., Gordon, M. A., Laskin, J. D., & Laskin, D. L. (2002). Regulation of cyclooxygenase-2 by nitric oxide in activated hepatic macrophages during acute endotoxemia. Journal of Leukocyte Biology, 71(6), 1005-1011. PMid:12050186.). PGE2 has been incriminated as an important mediator in the processes of inflammation (Ahmad et al., 2002Ahmad, N., Chen, L. C., Gordon, M. A., Laskin, J. D., & Laskin, D. L. (2002). Regulation of cyclooxygenase-2 by nitric oxide in activated hepatic macrophages during acute endotoxemia. Journal of Leukocyte Biology, 71(6), 1005-1011. PMid:12050186.). As previously reported, NO production is closely related to generation of PGE2 (Heo et al., 2010Heo, S. J., Yoon, W. J., Kim, K. N., Ahn, G. N., Kang, S. M., Kang, D. H., Affan, A., Oh, C., Jung, W. K., & Jeon, Y. J. (2010). Evaluation of anti-inflammatory effect of fucoxanthin isolated from brown algae in lipopolysaccharide-stimulated RAW 264.7 macrophages. Food and Chemical Toxicology, 48(8-9), 2045-2051. http://dx.doi.org/10.1016/j.fct.2010.05.003. PMid:20457205.
http://dx.doi.org/10.1016/j.fct.2010.05....
). Thus, chemo preventative agents that attenuate COX-2-mediated PGE2 production have had a healing effect in many inflammation-related diseases (Höcherl et al., 2002Höcherl, K., Dreher, F., Kurtz, A., & Bucher, M. (2002). Cyclooxygenase-2 inhibition attenuates lipopolysaccharide-induced cardiovascular failure. Hypertension, 40(6), 947-953. http://dx.doi.org/10.1161/01.HYP.0000041221.13644.B9. PMid:12468584.
http://dx.doi.org/10.1161/01.HYP.0000041...
). The results demonstrated that WEDII and WEDV decreased NO and PGE2 production in an LPS-stimulated RAW 264.7 macrophage incubation system by decreasing the expression of iNOS and COX-2.
3.5 Effect of the WED and WED fractions on TNF-α and IL-1β production in LPS-stimulated RAW 264.7 cells
LPS stimulated tremendous production of TNF-α. The WED and WED fractions were significantly suppressed production of TNF-α in the LPS-stimulated RAW 264.7 cells (Figure 5A). WEDV was the most active in suppressing TNF-α, showing no practical difference compared to that in LPS-untreated RAW 264.7 cells followed by WEDII.

The effect of WED and WED fractions on LPS-induced TNF-α (A) and IL-1β (B) production in RAW 264.7 cells. RAW 264.7 cells were prepared with WED and WED fractions for 1 h before being cultivated with 1 μg/mL LPS for 24 h. The supernatants were subsequently separated, and the amount of TNF-α a and IL-1β production was measured using an ELISA kit. Values are mean±S.D. (n=3). *a-eBars with different letters differ significantly at p<0.05 by Tukey test.
Similar tendencies in inhibition ability of WED and WED fractions on IL-1β production of LPS-stimulated RAW 264.7 cells were found (Figure 5B). WEDV with a concentration of 100 μg/mL most effectively showed suppression of IL-1β production.
Cytokines are proteins of low-molecular weight that influence immune cell function and are related during for activation of the inflammatory cascade (Kim et al., 2008Kim, J. Y., Kim, T. H., & Kim, S. S. (2008). Anti-inflammatory effect of a human prothrombinfragment-2-derived peptide, NSA9, in EOC microglia. Biochemical and Biophysical Research Communications, 368(3), 779-785. http://dx.doi.org/10.1016/j.bbrc.2008.01.142. PMid:18261978.
http://dx.doi.org/10.1016/j.bbrc.2008.01...
). TNF-α is a strong activator of macrophages and can stimulate the production and expression of IL-1β, PGE2, collagenase, and adhesion molecules. It elicits a number of physiological effects, including septic shock, inflammation, and cytotoxicity (Kim et al., 2010Kim, K. N., Heo, S. J., Yoon, W. J., Kang, S. M., Ahn, G., Yi, T. H., & Jeon, Y. J. (2010). Fucoxanthin inhibits the inflammatory response by suppressing the activation of NF-κB and MAPKs in lipopolysaccharide-induced RAW 264.7 macrophages. European Journal of Pharmacology, 649(1-3), 369-375. http://dx.doi.org/10.1016/j.ejphar.2010.09.032. PMid:20868674.
http://dx.doi.org/10.1016/j.ejphar.2010....
). IL-1β is considered as a central pro-inflammatory cytokine, mainly released by macrophages, and it acts a major part in the pathophysiology of rheumatoid arthritis (Lee et al., 2012bLee, S. J., Kim, E. K., Kim, Y. S., Hwang, J. W., Lee, K. H., Choi, D. K., Kang, H., Moon, S. H., Jeon, B. T., & Park, P. J. (2012b). Purification and characterization of a nitric oxide inhibitory peptide from Ruditapes philippinarum. Food and Chemical Toxicology, 50(5), 1660-1666. http://dx.doi.org/10.1016/j.fct.2012.02.021. PMid:22386812.
http://dx.doi.org/10.1016/j.fct.2012.02....
). Inflammatory stimuli, such as LPS, induce cytokines in the process of macrophage activation, which mediates tissue response in different phases of inflammation (Hseu et al., 2005Hseu, Y. C., Wu, F. Y., Wu, J. J., Chen, J. Y., Chang, W. H., Lu, F. J., Lai, Y. C., & Yang, H. L. (2005). Anti-inflammatory potential of Antrodia Camphorata through inhibition of iNOS, COX-2 and cytokines via the NF-kappaB pathway. International Immunopharmacology, 5(13-14), 1914-1925. http://dx.doi.org/10.1016/j.intimp.2005.06.013. PMid:16275626.
http://dx.doi.org/10.1016/j.intimp.2005....
). Because of their major parts in inflammation related reaction, down-regulation of these pro-inflammatory mediators is of the greatest importance in anti-inflammatory treatment. Here, we showed that WEDII and WEDV inhibited the LPS-stimulated production of TNF-α and IL-1β.
Soybean-derived peptides have shown ability in inhibiting pro-inflammatory mediators (Mejia & Dia, 2009Mejia, E. G., & Dia, V. P. (2009). Lunasin and lunasin-like peptides inhibit inflammation through suppression of NF-κB pathway in the macrophage. Peptides, 30(12), 2388-2398. http://dx.doi.org/10.1016/j.peptides.2009.08.005. PMid:19682518.
http://dx.doi.org/10.1016/j.peptides.200...
). Lunasin and lunasin-like peptides (5, 8 and 14 kDa) decreased both the production of NO and the expression of iNOS in LPS-stimulated RAW 264.7 macrophages. The efficacy of the three peptides in inhibiting the COX-2/PGE2 pathway follows the similar trend as in inhibiting the iNOS/NO pathway. These peptides described that the 5 kDa peptide has amino acid sequences consistent to the primary structure of soybean derived lunasin, the 8 kDa peptide showed partial sequence homology with 2S soy albumin, a lunasin precursor, and the 14 kDa peptide showed amino acid sequences, some of which corresponded to lunasin, among other proteins (Hernández-Ledesma et al., 2009Hernández-Ledesma, B., Hsieh, C. C., & de Lumen, B. O. (2009). Antioxidant and anti-inflammatory properties of cancer preventive peptide lunasin in RAW 264.7 macrophages. Biochemical and Biophysical Research Communications, 390(3), 803-808. http://dx.doi.org/10.1016/j.bbrc.2009.10.053. PMid:19836349.
http://dx.doi.org/10.1016/j.bbrc.2009.10...
). Thus, it can be proposed that WEDII and WEDV may contain peptides of a specific amino acid sequence representing anti-inflammatory activity.
Activated macrophages produce large quantities of NO and pro-inflammatory cytokines, which are involved in up-regulating inflammatory reactions. However, the overproduction of these mediators leads to in many acute and chronic human-related diseases, including hemorrhagic shock, including septic shock, rheumatoid arthritis, and multiple sclerosis (Bertolini et al., 2001Bertolini, A., Ottani, A., & Sandrini, M. (2001). Dual acting anti-inflammatory drugs: A reappraisal. Pharmacological Research, 44(6), 437-450. http://dx.doi.org/10.1006/phrs.2001.0872. PMid:11735348.
http://dx.doi.org/10.1006/phrs.2001.0872...
). Therefore, inhibiting these inflammatory mediators may be potentially effective for preventing inflammatory reactions and diseases. WEDII and WEDV effectively inhibited the production of pro-inflammatory cytokines; thus, it may be useful for preventing inflammatory related diseases.
4 Conclusion
Fractions of molecular weight range between 1 and 3 and 10 and 30 kDa (WEDII and WEDV) performed as an anti-inflammatory agent in macrophages via inhibition of NO/iNOS and PGE2/COX-2 pathways similarly suppression of the production of pro-inflammatory cytokines, including IL-1β and TNF-α. WEDV, in particular, showed the highest anti-inflammatory effect in LPS-induced production of inflammatory mediators. Many studies have reported that bioactive peptides derived from food protein possess a wide range of molecular weights. Therefore, each fractions of water extracts of protein hydrolysates can be a useful source of specific biological functions, if isolated, as well as doenjang.
Acknowledgements
This work was supported by a grant from Research year of Inje University in 2016 (20160008).
-
Practical Application: Water extracts of doenjang showed the highest anti-inflammatory effect in LPS-induced production of inflammatory mediators.
References
- Ahmad, N., Chen, L. C., Gordon, M. A., Laskin, J. D., & Laskin, D. L. (2002). Regulation of cyclooxygenase-2 by nitric oxide in activated hepatic macrophages during acute endotoxemia. Journal of Leukocyte Biology, 71(6), 1005-1011. PMid:12050186.
- Bertolini, A., Ottani, A., & Sandrini, M. (2001). Dual acting anti-inflammatory drugs: A reappraisal. Pharmacological Research, 44(6), 437-450. http://dx.doi.org/10.1006/phrs.2001.0872 PMid:11735348.
» http://dx.doi.org/10.1006/phrs.2001.0872 - Borek, C. (1991). Free radical processes in multistage carcinogenesis. Free Radical Research Communications, 13(1), 745-750. http://dx.doi.org/10.3109/10715769109145854 PMid:2060846.
» http://dx.doi.org/10.3109/10715769109145854 - Bowie, A., & O’Neill, L. A. (2000). Oxidative stress and nuclear factor-κB activation: a reassessment of the evidence in the light of recent discoveries. Biochemical Pharmacology, 59(1), 13-23. http://dx.doi.org/10.1016/S0006-2952(99)00296-8 PMid:10605930.
» http://dx.doi.org/10.1016/S0006-2952(99)00296-8 - Cam, A., & Mejia, E. G. (2012). RGD-peptide lunasin inhibits Akt-mediated NF-κB activation in human macrophages through interaction with the αVβ3 integrin. Molecular Nutrition & Food Research, 56(10), 1569-1581. http://dx.doi.org/10.1002/mnfr.201200301 PMid:22945510.
» http://dx.doi.org/10.1002/mnfr.201200301 - Choi, M. S., & Rhee, K. C. (2006). Production and Processing of soybeans and nutrition and safety of isoflavone and other soy products for human health. Journal of Medicinal Food, 9(1), 1-10. http://dx.doi.org/10.1089/jmf.2006.9.1 PMid:16579721.
» http://dx.doi.org/10.1089/jmf.2006.9.1 - Gibbs, B. F., Zougman, A., Masse, R., & Mulligan, C. (2004). Production and characterization of bioactive peptides from soy hydrolysate and soy fermented food. Food Research International, 37(2), 123-131. http://dx.doi.org/10.1016/j.foodres.2003.09.010
» http://dx.doi.org/10.1016/j.foodres.2003.09.010 - Heo, S. J., Yoon, W. J., Kim, K. N., Ahn, G. N., Kang, S. M., Kang, D. H., Affan, A., Oh, C., Jung, W. K., & Jeon, Y. J. (2010). Evaluation of anti-inflammatory effect of fucoxanthin isolated from brown algae in lipopolysaccharide-stimulated RAW 264.7 macrophages. Food and Chemical Toxicology, 48(8-9), 2045-2051. http://dx.doi.org/10.1016/j.fct.2010.05.003 PMid:20457205.
» http://dx.doi.org/10.1016/j.fct.2010.05.003 - Hernández-Ledesma, B., Hsieh, C. C., & de Lumen, B. O. (2009). Antioxidant and anti-inflammatory properties of cancer preventive peptide lunasin in RAW 264.7 macrophages. Biochemical and Biophysical Research Communications, 390(3), 803-808. http://dx.doi.org/10.1016/j.bbrc.2009.10.053 PMid:19836349.
» http://dx.doi.org/10.1016/j.bbrc.2009.10.053 - Höcherl, K., Dreher, F., Kurtz, A., & Bucher, M. (2002). Cyclooxygenase-2 inhibition attenuates lipopolysaccharide-induced cardiovascular failure. Hypertension, 40(6), 947-953. http://dx.doi.org/10.1161/01.HYP.0000041221.13644.B9 PMid:12468584.
» http://dx.doi.org/10.1161/01.HYP.0000041221.13644.B9 - Hou, Y., Liu, W., Cheng, Y., Zhou, J., Wu, L., & Yang, G. (2014). Production optimization and characterization of immunomodulatory peptides obtained from fermented goat placenta. Food Science and Technology, 34(4), 723-729. http://dx.doi.org/10.1590/1678-457X.6448
» http://dx.doi.org/10.1590/1678-457X.6448 - Hseu, Y. C., Wu, F. Y., Wu, J. J., Chen, J. Y., Chang, W. H., Lu, F. J., Lai, Y. C., & Yang, H. L. (2005). Anti-inflammatory potential of Antrodia Camphorata through inhibition of iNOS, COX-2 and cytokines via the NF-kappaB pathway. International Immunopharmacology, 5(13-14), 1914-1925. http://dx.doi.org/10.1016/j.intimp.2005.06.013 PMid:16275626.
» http://dx.doi.org/10.1016/j.intimp.2005.06.013 - Hwang, J. S., Yoo, H. J., Song, H. J., Kim, K. K., Chun, Y. J., Matsui, T., & Kim, H. B. (2011). Inflammation related signaling pathways implicating TGF𝛽 are revealed in the expression profiling of MCF7 cell treated with fermented soybean, Chungkookjang. Nutrition and Cancer, 63(4), 645-652. http://dx.doi.org/10.1080/01635581.2011.551987 PMid:21547849.
» http://dx.doi.org/10.1080/01635581.2011.551987 - Kalyanaraman, B., Darley-Usmar, V., Davies, K. J., Dennery, P. A., Forman, H. J., Grisham, M. B., Mann, G. E., Moore, K., Roberts, L. J. 2nd, & Ischiropoulos, H. (2012). Measuring reactive oxygen and nitrogen species with fluorescent probes: challenges and limitations. Free radical biology and medicine, 52(1), 1-6. http://dx.doi.org/10.1016/j.freeradbiomed.2011.09.030
» http://dx.doi.org/10.1016/j.freeradbiomed.2011.09.030 - Kim, J. Y., Kim, T. H., & Kim, S. S. (2008). Anti-inflammatory effect of a human prothrombinfragment-2-derived peptide, NSA9, in EOC microglia. Biochemical and Biophysical Research Communications, 368(3), 779-785. http://dx.doi.org/10.1016/j.bbrc.2008.01.142 PMid:18261978.
» http://dx.doi.org/10.1016/j.bbrc.2008.01.142 - Kim, K. N., Heo, S. J., Yoon, W. J., Kang, S. M., Ahn, G., Yi, T. H., & Jeon, Y. J. (2010). Fucoxanthin inhibits the inflammatory response by suppressing the activation of NF-κB and MAPKs in lipopolysaccharide-induced RAW 264.7 macrophages. European Journal of Pharmacology, 649(1-3), 369-375. http://dx.doi.org/10.1016/j.ejphar.2010.09.032 PMid:20868674.
» http://dx.doi.org/10.1016/j.ejphar.2010.09.032 - Kitts, D. D., & Weiler, K. (2003). Bioactive proteins and peptides from food sources. Applications of bioprocesses used in isolation and recovery. Current Pharmaceutical Design, 9(16), 1309-1323. http://dx.doi.org/10.2174/1381612033454883 PMid:12769739.
» http://dx.doi.org/10.2174/1381612033454883 - Ko, S. C., & Jeon, Y. J. (2015). Anti-inflammatory effect of enzymatic hydrolysates from Styela clava flesh tissue in lipopolysaccharide-stimulated RAW 264.7 macrophages and in vivo zebrafish model. Nutrition Research and Practice, 9(3), 219-226. http://dx.doi.org/10.4162/nrp.2015.9.3.219 PMid:26060532.
» http://dx.doi.org/10.4162/nrp.2015.9.3.219 - Korhonen, H., & Pihlanto, A. (2003). Food-derived bioactive peptides-opportunities for designing future foods. Current Pharmaceutical Design, 9(16), 1297-1308. http://dx.doi.org/10.2174/1381612033454892 PMid:12769738.
» http://dx.doi.org/10.2174/1381612033454892 - Lee, H. S., Ryu, D. S., Lee, G. S., & Lee, D. S. (2012a). Anti-inflammatory effects of dichloromethane fraction from Orostachys japonicus in RAW 264.7 cells: suppression of NF-κB activation and MAPK signaling. Journal of Ethnopharmacology, 140(2), 271-276. http://dx.doi.org/10.1016/j.jep.2012.01.016 PMid:22285522.
» http://dx.doi.org/10.1016/j.jep.2012.01.016 - Lee, S. H. (2012). Physiological activities of water extracts of doenjang fractionated to their molecular weight of nitrogenous compounds (Ph.D. Dissertation). Inje University, Gimhae, Gyeongnam, Korea.
- Lee, S. H., Jung, J. H., Lee, H. A., Lee, H.-S., Jeong, H. D., & Cherl, J. (2014). Fractionation of nitrogenous compounds from water extracts of doenjang and their physiological activities. Food Science and Biotechnology, 23(2), 523-529. http://dx.doi.org/10.1007/s10068-014-0072-x
» http://dx.doi.org/10.1007/s10068-014-0072-x - Lee, S. J., Kim, E. K., Kim, Y. S., Hwang, J. W., Lee, K. H., Choi, D. K., Kang, H., Moon, S. H., Jeon, B. T., & Park, P. J. (2012b). Purification and characterization of a nitric oxide inhibitory peptide from Ruditapes philippinarum. Food and Chemical Toxicology, 50(5), 1660-1666. http://dx.doi.org/10.1016/j.fct.2012.02.021 PMid:22386812.
» http://dx.doi.org/10.1016/j.fct.2012.02.021 - Lee, S. O., Kim, S. Y., Han, S. M., Kim, H. M., Ham, S. S., & Kang, I. J. (2006). Corni fructus scavengers hydroxyl radicals and decreases oxidative stress in Endothelial cells. Journal of Medicinal Food, 9(4), 594-598. http://dx.doi.org/10.1089/jmf.2006.9.594 PMid:17201652.
» http://dx.doi.org/10.1089/jmf.2006.9.594 - Lumen, B. O. (2005). Lunasin: A cancer preventive peptide. Nutrition Reviews, 63(1), 16-21. http://dx.doi.org/10.1111/j.1753-4887.2005.tb00106.x PMid:15730231.
» http://dx.doi.org/10.1111/j.1753-4887.2005.tb00106.x - Mallikarjun Gouda, K. G., Gowda, L. R., Rao, A. G., & Prakash, V. (2006). Angiotensin I-converting enzyme inhibitory peptide derived from glycinin, the 11S globulin of soybean (Glycine max). Journal of Agricultural and Food Chemistry, 54(13), 4568-4573. http://dx.doi.org/10.1021/jf060264q PMid:16786999.
» http://dx.doi.org/10.1021/jf060264q - Mejia, E. G., & Dia, V. P. (2009). Lunasin and lunasin-like peptides inhibit inflammation through suppression of NF-κB pathway in the macrophage. Peptides, 30(12), 2388-2398. http://dx.doi.org/10.1016/j.peptides.2009.08.005 PMid:19682518.
» http://dx.doi.org/10.1016/j.peptides.2009.08.005 - Murakami, A., & Ohigashi, H. (2007). Targeting NOX, INOS and COX-2 in inflammatory cells: chemoprevention using food phytochemicals. International Journal of Cancer, 121(11), 2357-2363. http://dx.doi.org/10.1002/ijc.23161 PMid:17893865.
» http://dx.doi.org/10.1002/ijc.23161 - Tsuruki, T., Kishi, K., Takahashi, M., Tanaka, M., Matsukawa, T., & Yoshikawa, M. (2003). Soymetide, an immunostimulating peptide derived from soybean β-conglycinin, is an fMLP agonist. FEBS Letters, 540(1-3), 206-210. http://dx.doi.org/10.1016/S0014-5793(03)00265-5 PMid:12681509.
» http://dx.doi.org/10.1016/S0014-5793(03)00265-5 - Vernaza, M. G., Dia, V. P., de Mejia, E. G., & Chang, Y. K. (2012). Antioxidant and antiinflammatory properties of germinated and hydrolysed Brazilian soybean flours. Food Chemistry, 134(4), 2217-2225. http://dx.doi.org/10.1016/j.foodchem.2012.04.037 PMid:23442677.
» http://dx.doi.org/10.1016/j.foodchem.2012.04.037 - Wongputtisin, P., Khanongnuch, C., Pongpiachan, P., & Lumyong, S. (2007). Antioxidant activity improvement of soybean meal by microbial fermentation. Research Journal of Microbiology, 2(7), 577-583. http://dx.doi.org/10.3923/jm.2007.577.583
» http://dx.doi.org/10.3923/jm.2007.577.583 - Yoshikawa, M., Fujita, H., Matoba, N., Takenaka, Y., Yamamoto, T., Yamauchi, R., Tsuruki, H., & Takahata, K. (2000). Bioactive peptides derived from food proteins preventing lifestyle-related diseases. BioFactors, 12(1-4), 143-146. http://dx.doi.org/10.1002/biof.5520120122 PMid:11216476.
» http://dx.doi.org/10.1002/biof.5520120122
Publication Dates
-
Publication in this collection
04 July 2019 -
Date of issue
Oct-Dec 2019
History
-
Received
04 May 2018 -
Accepted
31 Mar 2019





