Abstract
OBJECTIVE: To review the literature on the interaction between sleep and the immune system. METHOD: A search on Web of Science and Pubmed database including the keywords sleep, sleep deprivation, stress, hypothalamic-pituitary-adrenal axis, immune system, and autoimmune diseases. RESULTS: On Web of Science, 588 publications were retrieved; 61 references, more significant and closer to our objective, were used, including original articles and review papers. CONCLUSION: Sleep deprivation and immune system exert a bidirectional influence on each other. Since sleep deprivation is considered a stressor, inasmuch as it induces elevation of cortisol or corticosterone levels in humans and rodents, respectively, and given the well-known immunosuppressive effect of glucocorticoids, we propose that increased activation of the hypothalamic-pituitary-adrenal axis is a major mediator of the immune alterations observed in patients with insomnia or in sleep deprived subjects.
Sleep deprivation; Stress; Hypothalamic-pituitary-adrenal axis; Immune system; Autoimmune diseases
Resumo
OBJETIVO: Revisar a literatura a respeito da interação entre sono e sistema imunológico. MÉTODO: Busca no Web of Science e no PubMed com os descritores: sono, privação de sono, estresse, eixo hipotálamo-pituitária-adrenal, sistema imunológico e doenças auto-imunes. RESULTADOS: Foram encontrados 588 artigos no Web of Science. As 61 referências mais significativas e mais relacionadas aos objetivos do estudo foram utilizadas. Foram incluídos artigos originais e de revisão. CONCLUSÃO: A privação de sono e o sistema imunológico exercem e sofrem influências mútuas. A privação de sono é considerada um estressor, uma vez que induz a elevação do cortisol em seres humanos - ou da corticosterona em roedores. Os glicocorticóides, por sua vez, exercem um efeito imunossupressor. Por essas razões, foi proposto que o aumento da ativação do eixo hipotálamo-pituitária-adrenal seja um importante mediador das alterações imunológicas observadas em pacientes com insônia ou privados de sono.
Privação de sono; Estresse; Eixo hipotálamo-pituitária-adrenal; Sistema imunológico; Doenças auto-imunes
ARTICLES
Immune outcomes of sleep disorders: the hypothalamic-pituitary-adrenal axis as a modulatory factor
Beatriz Duarte Palma; Paula Ayako Tiba; Ricardo Borges Machado; Sergio Tufik; Deborah Suchecki
Department of Psychobiology, Universidade Federal de São Paulo (UNIFESP), São Paulo (SP), Brazil
ABSTRACT
OBJECTIVE: To review the literature on the interaction between sleep and the immune system.
METHOD: A search on Web of Science and Pubmed database including the keywords sleep, sleep deprivation, stress, hypothalamic-pituitary-adrenal axis, immune system, and autoimmune diseases.
RESULTS: On Web of Science, 588 publications were retrieved; 61 references, more significant and closer to our objective, were used, including original articles and review papers.
CONCLUSION: Sleep deprivation and immune system exert a bidirectional influence on each other. Since sleep deprivation is considered a stressor, inasmuch as it induces elevation of cortisol or corticosterone levels in humans and rodents, respectively, and given the well-known immunosuppressive effect of glucocorticoids, we propose that increased activation of the hypothalamic-pituitary-adrenal axis is a major mediator of the immune alterations observed in patients with insomnia or in sleep deprived subjects.
Descriptors: Sleep deprivation; Stress; Hypothalamic-pituitary-adrenal axis; Immune system; Autoimmune diseases
Introduction
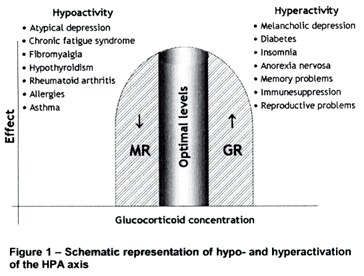
The mammalian organism is gifted with a system whose main function is to maintain the inner balance. This neuroendocrine system, known as the hypothalamic-pituitary-adrenal (HPA) axis, is mobilized every time there is a threat to life, regardless of whether it is real or imaginary, so that appropriate behaviors will be displayed and metabolic changes will take place to provide the energy necessary to deal with such situations. Inappropriate, either hypo- or hyperactivation of the HPA axis will lead to illness (Figure 1), including sleep- and immune-related disorders. Therefore, we propose that the HPA axis could mediate the relationship between sleep disorders, mainly insomnia, and the immune system.
Human sleep
The sleep in human beings is generally divided in two main phases: Rapid Eye Movement (REM) sleep and Non-REM sleep. Non-REM sleep can be subdivided in four phases: stages 1 and 2, and stages 3 and 4, which, together, are known as delta sleep. As shown in Figure 2, the stages follow a sequence in which there is a predominance of delta sleep during the first half of the night and longer and more frequent events of REM sleep during the last third of the night.
Sleep evolves in cycles of NREM and REM phases, which, together, take approximately 90 minutes. During these cycles, specific physiological events take place, such as release of growth hormone during delta sleep, whereas the lowest activity of the HPA axis is observed during delta sleep.1 Cytokines show a nocturnal profile similar to that found for the sleep-dependent hormone, the growth hormone. Sleep onset is associated with an increase in circulating levels of some cytokines, with peak values occurring 2.5 hours after sleep onset.2 The circadian rhythm of pro-inflammatory cytokines may explain, at least in part, the nighttime exacerbations of immunoinflammatory disorders such as asthma and rheumatoid arthritis.
The HPA axis and the stress response
The secretion of corticotrophin-releasing factor (CRF), adrenocorticotropin hormone (ACTH), cortisol or corticosterone (CORT), noradrenaline (NA), and adrenaline is also related to wake-sleep cycle, but is primordially controlled by circadian rhythm. Thus, peak activity of both HPA axis and sympathetic autonomic nervous system occur shortly before the active period (daytime for human beings and nighttime for rodents), and minimum activity is observed shortly before sleep1 (Figure 3).
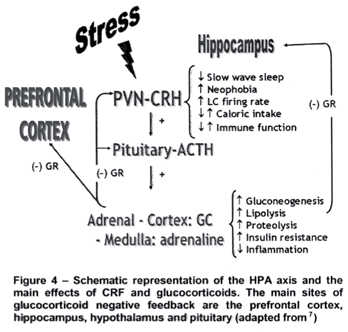
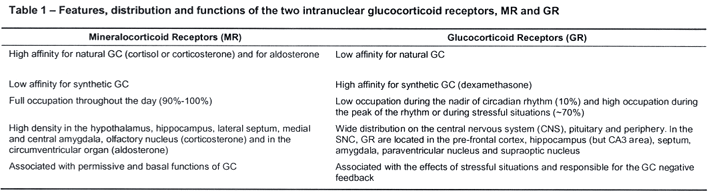
Therefore, the activity of the HPA axis is increased in preparation to deal with stress and in response to actual stressful events. Thus, it is vital that the system is activated when necessary, but even more so, that the system gets shut down as soon as the situation is dealt with. The most important regulator of this system is its end product, the glucocorticoid cortisol (in primates and human beings) or corticosterone (in rodents). In a classical review paper, Munck et al. have proposed that "the physiological function of stress-induced increases in glucocorticoid levels is to protect not against the source of stress itself, but against the normal defence reactions that are activated by stress. Glucocorticoids accomplish this function by turning off those defence reactions, thus preventing them from overshooting and themselves threatening homeostasis".3 This means that if unchecked by the glucocorticoid negative feedback system, the activation of the HPA axis may itself become detrimental. Glucocorticoids (GCs), in fact, exhibit permissive and suppressive effects that are complementary, for the former prepare or prime the defence mechanisms for action and the latter limit these actions, by binding to type II or glucocorticoid receptors (GR) to exert this negative feedback action (Figure 4). GR are ubiquitously distributed in the CNS and periphery, including the immune system, whereas type I or mineralocorticoid receptors (MR) show a more restricted distribution, being located in the limbic system, hippocampus and septum (Table 1). Behavioral and neuroendocrine studies suggest that MR are involved with the permissive and/or circadian actions of GCs, whereas GR are responsible for the suppressive effects, restoring the disturbed homeostasis resulting from stress exposure.4-6,8
Hormones of the HPA axis and sleep
Classically, activation of the HPA and/or the sympathetic nervous systems results in waking. In human beings or in animals, these hormones are associated with attention, vigilance and arousal. This plethora of effects is intuitive for nobody will feel asleep in face of danger or a life threat.
As mentioned above, the lowest levels of cortisol are obtained during delta sleep. This might suggest that cortisol release is inhibited by delta sleep, as had been believed for a long time.9 However, a recent study has demonstrated that, contrary to previous belief, is the lack of cortisol pulse that allows delta sleep to occur, since temporally, lack of cortisol pulse precedes delta sleep.10
Another interesting aspect refers to changes in the sensitivity of the axis throughout the sleep cycle, inasmuch as pituitary response to a corticotrophin-releasing factor/arginine vasopressin (CRF/AVP) challenge in healthy volunteers kept awake is higher than when they are allowed to sleep, regardless of the time of sleep deprivation,11 suggesting that reduced response of the pituitary to CRF is not merely a reflex of circadian influences, but it is rather inhibited by sleep onset.12
Numerous findings have confirmed the effects of CRF on sleep. In animals, intra LC injection of CRF stimulates the firing rate of these neurons,13 which are known to be more active during waking.14 Moreover, i.c.v. administration of low doses of CRF results in EEG waking pattern, whereas blockade of CRF receptors or inactivation of CRF synthesis reduces spontaneous waking and increases delta sleep in rats treated during their active period.15-16
Additional evidence for the role of CRF on the regulation of waking is given by Lewis rats, which present a genetic deficiency in CRF production. Therefore, these rats show a hypoactive HPA axis and, consequently, they also spend less time in waking and more time in slow wave sleep than rats with a normal HPA axis activity.17 Blockade of cortisol synthesis with metyrapone, an inhibitor of the 11b-hydroxilase, leads to augmented activity of the CRF system (due to a chemical removal of the negative feedback system) and, as a result, reduction of delta sleep.18
Administration of CRF to human beings also impairs sleep, but the consequences also appear to be age-dependent. Whilst young men (20-28 years old) only exhibit increased waking time during the second half of the night by virtue of reduced REM sleep, middle aged volunteers (37-54 years old) display more severe impairments, including augmented waking time after sleep onset, throughout the entire night.19
In regards to the effects of GCs on sleep, data are scarce and sometimes difficult to interpret due to the close relationship among the components of the HPA axis. Therefore, high levels of GCs might lead to direct outcomes, which could also be the result of CRF-dependent regulation. For instance, repeated administration of hydrocortisone increases theta and delta frequencies (synchronized activity at the EEG), indicating increased delta sleep. On the contrary, REM sleep is reduced. These effects may be caused by an inhibition of CRF actions on sleep.20
However, it must be emphasized that optimum levels of cortisol are necessary for good quality sleep. For instance, in rats, adrenalectomy leads to augmented slow wave sleep, but reduces REM sleep.21-22 Likewise, patients with adrenal insufficiency due to Addison's Disease present impaired REM sleep, and treatment with hydrocortisone normalizes these disturbances, increasing the percentage and reducing the latency to REM sleep.23
Patients under chronic treatment with methylprednisolone (a GR agonist) exhibit reduction of REM sleep latency, augmented REM sleep time, and a dislocation of slow-wave sleep (SWS) from the first to the second half of the sleep period. In contrast, the MR antagonist canrenoate only reduces SWS.24-25 Mifepristone, a mixed GR progesterone receptor antagonist, diminishes the SWS and REM time with increase of sleep latency. The combination of dexamethasone and spironolactone, a MR antagonist, reduces REM sleep; and dexamethasone followed by mifepristone diminishes total sleep time (SWS and REM) and increases the arousals during the night.26 In respect to the weak influence of MR receptors on sleep control, another study has shown that the MR agonists deoxycorticosterone and spironolactone do not affect sleep EEG substantially in human beings.27
The results presented above suggest that corticosteroid-induced changes in SWS are mediated via MR-like central receptors in humans, whereas changes in REM sleep involve GR-mediated events.
The immune system and the HPA axis
GCs represent the most powerful endogenous anti-inflammatory and immunosuppressive agent, interfering with virtually every step of the immunoinflammatory responses. Because GCs are regarded as the most effective anti-inflammatory drugs, they were considered, for many years, as purely immunosuppressant agents. Today, the immunomodulatory role of the GCs is well recognized, such as modulating T and B-cell maturation, selection, proliferation and influence on the trafficking, and activation of inflammatory cells by inhibiting the expression of adhesion molecules and major histocompatibility complex (MHC) class II antigens in leukocytes. In addition, GCs inhibit the production of a wide range of cytokines including IL-1, IL-2, IL-6 and TNF-a and decrease the effects of certain inflammatory molecules on various target tissues. At physiological concentrations, GCs inhibit T helper-type I (Th1) and enhance Type 2 (Th2) cytokine production, thus shifting immune responses from Th1 to a Th2 pattern.28 Th1 and Th2 responses are defined by the different types of cytokines secreted.29 For instance, Th1 response is defined by secretion of pro-inflammatory cytokines, whereas Th2 secretes the major anti-inflammatory cytokines. Both responses play an important role in the course of an immune response, insofar as Th1 response promotes cellular immunity, whist the Th2 initiates humoral immunity and counteracts the Th1 response. The Th1/Th2 unbalance is a crucial factor that determines the manifestation of some diseases such as autoimmune and inflammatory ones. Some results strongly suggest that a high reactivity of the HPA axis is associated with a high capacity to generate Th2-type responses, whereas low HPA axis reactivity goes with a higher capacity to generate Th1-type response.30
The HPA axis-immune system relationship exhibits a bimodal pattern in which GCs may function as either an inhibitory or permissive factor depending on its circulating levels. Disturbances at any level of the HPA axis or on the action of GCs results in immune system imbalance, which consequently is harmful to the organism. GCs act as modulators of the immune system, for either excess or deficient release are associated with illness. On one hand, hyper-stimulation of the HPA axis, with excessive secretion of GCs, results in intense immunosuppression and increased susceptibility to infection; on the other hand, insufficient secretion of GCs leads to inflammatory and autoimmune diseases.31
Let us consider, for instance, a situation in which the stressful stimulus is an injury to the skin. Appropriate stress response, with adequate levels of cortisol and adrenaline, results in vasoconstriction to prevent hemorrhage, increased traffic of leukocytes to fight any possible antigen and increased production of fibrinogen to help wound healing.32 CRF also exhibits this bimodal pattern of effect on the immune system, since it decreases T cell proliferation and natural killer (NK) cell cytotoxicity, but can also enhance B cell proliferation and the proliferative lymphocyte response to various mitogens.8 Therefore, in an initial demand, these hormones are immunostimulant, rather than immunosuppressive. The role of GCs in establishing homeostasis includes their immunosuppressive effect, for it can reduce the likelihood of autoimmune overshoot. Polyclonal responses are induced by immune challenges, which may increase the risk of autoimmunity. Under physiological conditions GCs are selective, "sculpting" the immune response so that superfluous or autoimmune-pron e components are selectively inhibited, due to the preferential targeting by GCs of lymphocytes that are less active or that produce antibodies with lower affinities for the antigen.8
The use of animal models has led to many important findings highlighting how essential the stress response is for both the physiological regulation of the immune system and for the development and manifestation of inflammatory and autoimmune diseases. The rat strains Lewis (LEW/N) and Fischer (F344/N) are widely used to assess neuroendocrine regulation of many aspects of autoimmunity. LEW/N rats are more susceptible to the development of autoimmune/inflammatory disorders in response to antigenic stimuli, since this strain exhibits a hyporesponsiveness of the HPA axis. The F344/N strain, on the contrary, is relatively resistant to these disorders due to augmented activity of the HPA axis.33
Another extremely intriguing feature is the bidirectional relationship between the HPA axis and the immune system. Endogenous GCs production is tightly regulated with activation of the immune system, as part of the systemic stress response. Inflammatory and immune responses lead to a rapid activation of the HPA axis and production of GCs. The activation of the HPA axis was found to occur on the hypothalamic level34 and via the pituitary gland but some data have indicated that the adrenal may be a direct target for cytokine action during the immune response.35 Other cytokines, including IL-2, IL-6, tumor necrosis factor-a (TNF-a), and interferon-g (IFN-g), have been shown to stimulate the adrenocortical axis, although none with the potency of IL-1.8 The increased release of GCs in the course of immune challenges represents an important regulatory mechanism, providing a negative feedback in response to the overproduction of cytokines.
Inadequate sleep, the immune system and the HPA axis
The interaction between sleep and immune system has been systematically studied in the last decade. This bidirectional relationship is based on the lethargy and strong sleepiness that occur during infections and inflammatory conditions and on the augmented susceptibility to infections as a consequence of sleep deprivation.
An ever-growing number of studies have been performed in order to assess the impact of inadequate sleep on the quality of life, morbidity and mortality. For instance, a retrospective study conducted in Denmark has demonstrated that nighttime-working women showed augmented risk for breast cancer than daytime-working women.36 In an elegant in vitro study, the effects of cortisol alone or in association with melatonin on human lymphocyte proliferation after stimulation with concanavalin A (CON A) have been studied. Incubation with either cortisol alone (such as during the day) or with the combination of cortisol and melatonin (such as in shift workers) inhibits even further the lymphocyte proliferation, indicating that disruptions of the balance between cortisol and melatonin are detrimental to optimum activity of the immune system.37
Laboratory studies have shown that partial sleep deprivation following immunization against hepatitis A reduces the production of antibody titers approximately 4 weeks after immunization.38 In addition, partially sleep-deprived individuals also exhibit impairment of antibody production against Influenza virus. The antibody titers were determined in the fourth week postimmunization, at a time when production reaches a peak.39 These findings highlight the importance of ideal sleep, given that the effects of sleep deprivation can be manifested much later. These effects may be at least partly explained by the well-known stressful nature of sleep deprivation, which has been shown both in human beings40 and animals.41-42
Sleep disorders in which sleep deprivation is a major feature, such as insomnia and sleep apnea, result in major alterations in the immune system, which may be mediated by the augmented activity of the HPA axis and/or the sympathetic nervous system. Stress is considered one of the major triggering factors of insomnia, inasmuch as epidemiologic studies report that primary insomnia patients attribute the onset of the disorder to some stressful event.43-45 Chronic internalization of feelings may cause a psychological and physiological arousal, leading, ultimately, to insomnia. Some authors propose that increased activity of the HPA axis produces sleep fragmentation, which, in turn, elevates circulating levels of cortisol,46 whereas others43 believe in the existence of a mechanism of cortical hyperarousal in insomniac patients triggered by emotional, cognitive and physiological components. Therefore, the hyperactivity exacerbates vigilance and impacts negatively on sleep, thus forming a vicious circle in which difficulty to sleep becomes the stressful factor itself.47 This hypothesis has been confirmed by the positive correlation between elevated levels of cortisol, increased activity of the sympathetic system and time of waking in chronic insomniacs.48 However, there has always been a question of whether insomnia leads to increased HPA activity, or increased activity of the HPA axis is a risk factor for the development of stress-induced insomnia. It has been shown that in chronic insomniacs, plasma levels of ACTH and cortisol are elevated exactly during the nadir of the circadian rhythm, i.e., between 10 pm and 2 am, without any change in the circadian pattern of secretion but a flattening of the amplitude. The authors have interpreted these results as indicative that increased activity of the HPA axis is responsible for poor sleep in insomniacs, because if it were a response to bad sleep quality, ACTH and cortisol levels should be elevated in the morning as a response to the poor sleep.49
In regards to the correlation between immune alterations and insomnia, it has been shown that chronic insomnia patients exhibit a reduction of CD3+, CD4+ and CD8+ cells50 and a predominance of Th2 response, i.e., enhanced humoral immune response.51 It is worth emphasizing that alterations in Th1/Th2 balance are characteristic of autoimmune diseases. One of the major effects of GCs on the immune system is the suppression of cellular immunity and enhancement of humoral immunity, a process that is also driven by the Th1 to Th2 shift in cytokine profile production. Therefore, increased levels of cortisol could at least partly explain the change in Th1/Th2 response in insomniac patients.
Concluding remarks
Sleep is regulated by a myriad of neurotransmitters and hormones that work in concert to guarantee that such fundamental process is not severely disturbed by environmental factors. Likewise, a well-preserved immune system is essential for survival. Failure in maintaining these systems functioning in perfect harmony can be regarded as a stressor, thus leading to activation of the HPA axis, which can result in further disruptions. Therefore, the adequate stress response mounted by an individual is vital for the perfect functioning of these systems. Thus, although many other mediators play a role in sleep and the activity of the immune system, we believe that exaggerated secretion of GCs is the major mediator of sleep disorders-induced deficits in the immune system.
Acknowledgements
The authors' work is supported by Associação Fundo de Incentivo à Psicofarmacologia (AFIP) and Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP). Paula Ayako Tiba is the recipient of a Ph.D. scholarship from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) and Ricardo Borges Machado is the recipient of a Ph.D. scholarship from FAPESP. Deborah Suchecki and Sergio Tufik are recipients of fellowships from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
References
Financing: Associação Fundo de Incentivo à Psicofarmacologia (AFIP) and Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) - CEPID # 98/14303-3.
Conflict of interests: None
References
- 1 Born J, Fehm HL. The neuroendocrine recovery function of sleep. Noise Health 2000;2(7):25-38.
- 2. Haus E, Smolensky MH. Biologic rhythms in the immune system. Chronobiol Int 1999;16(5):581-622.
- 3. Munck A, Guyre PM, Holbrook NJ. Physiological functions of glucocorticoids in stress and their relation to pharmacological actions. Endocr Rev. 1984;5(1):25-44.
- 4. Dallman MF, Akana SF, Cascio CS, Darlington DN, Jacobson L, Levin N. Regulation of ACTH secretion: variations on a theme of B. Recent Prog Horm Res. 1987;43:113-73.
- 5. De Kloet ER, Reul JM. Feedback action and tonic influence of corticosteroids on brain function: a concept arising from the heterogeneity of brain receptor systems. Psychoneuroendocrinology. 1987;12(2):83-105.
- 6. Ratka A, Sutanto W, De Kloet ER. On the role of brain mineralocorticoid (Type I) and glucocorticoid (Type II) receptors in neuroendocrine regulation. Neuroendocrinology. 1989;50(2):117-23.
- 7. Arborelius L, Owens MJ, Plotsky PM, Nemeroff CB. The role of corticotropin-releasing factor in depression and anxiety disorders. J Endocrinol. 1999;160(1):1-12.
- 8. Sapolsky RM, Romero LM, Munck AU. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr Rev. 2000;21(1):55-89.
- 9. Weitzman ED, Zimmerman JC, Czeisler CA, Ronda J. Cortisol secretion is inhibited during sleep in normal man. J Clin Endocrinol Metab. 1983;56(2):352-8.
- 10. Gronfier C, Brandenberger G. Ultradian rhythms in pituitary and adrenal hormones: their relations to sleep. Sleep Med Rev. 1998;2(1):17-29.
- 11. Bierwolf C, Struve K, Marshall L, Born J, Fehm HL. Slow wave sleep drives inhibition of pituitary-adrenal secretion in humans. J Neuroendocrinol. 1997;9(6):479-84.
- 12. Spath-Schwalbe E, Uthgenannt D, Voget G, Kern W, Born J, Fehm HL. Corticotropin-releasing hormone-induced adrenocorticotropin and cortisol secretion depends on sleep and wakefulness. J Clin Endocrinol Metab. 1993;77(5):1170-3.
- 13. Curtis AL, Lechner SM, Paucovich LA, Valentino RJ. Activation of the locus coeruleus noradrenergic system by intracoerulear microinfusion of corticotropin-releasing factor: effects on discharge rate, cortical norepinephrine levels and cortical electroencephalographic activity. J Pharmacol Exp Ther. 1997;281(1):163-72.
- 14. Aston-Jones G, Bloom FE. Activity of norepinephrine-containing locus coeruleus neurons in behaving rats anticipates fluctuations in the sleep-waking cycle. J Neurosci. 1981;1(8):876-86.
- 15. Chang FC, Opp MR. Blockade of corticotropin-releasing hormone receptors reduces spontaneous waking in the rat. Am J Physiol. 1998;275(3 Pt 2):R793-802.
- 16. Chang FC, Opp MR. A corticotropin-releasing hormone antisense oligodeoxynucleotide reduces spontaneous waking in the rat. Regul Pept. 2004;117(1):43-52.
- 17. Opp MR. Rat strain differences suggest a role for corticotropin-releasing hormone in modulating sleep. Physiol Behav. 1998;63(1):67-74.
- 18. Jahn H, Kiefer F, Schick M, Yassouridis A, Steiger A, Kellner M, Wiedemann K. Sleep endocrine effects of the 11-beta-hydroxysteroiddehydrogenase inhibitor metyrapone. Sleep. 2003;26(7):823-9.
- 19. Vgontzas AN, Bixler EO, Wittman AM, Zachman K, Lin HM, Vela-Bueno A, Kales A, Chrousos GP. Middle-aged men show higher sensitivity of sleep to the arousing effects of corticotropin-releasing hormone than young men: clinical implications. J Clin Endocrinol Metab. 2001;86(4):1489-95.
- 20. Friess E, Tagaya H, Grethe C, Trachsel L, Holsboer F. Acute cortisol administration promotes sleep intensity in man. Neuropsychopharmacology. 2004;29(3):598-604.
- 21. Bradbury MJ, Dement WC, Edgar DM. Effects of adrenalectomy and subsequent corticosterone replacement on rat sleep state and EEG power spectra. Am J Physiol. 1998;275(2 Pt 2):R555-65.
- 22. Marinesco S, Bonnet C, Cespuglio R. Influence of stress duration on the sleep rebound induced by immobilization in the rat: a possible role for corticosterone. Neuroscience. 1999;92(3):921-33.
- 23. Garcia-Borreguero D, Wehr TA, Larrosa O, Granizo JJ, Hardwick D, Chrousos GP, Friedman TC. Glucocorticoid replacement is permissive for rapid eye movement sleep and sleep consolidation in patients with adrenal insufficiency. J Clin Endocrinol Metab. 2000;85(11):4201-6.
- 24. Born J, DeKloet ER, Wenz H, Kern W, Fehm HL. Gluco- and antimineralocorticoid effects on human sleep: a role of central corticosteroid receptors. Am J Physiol. 1991;260 (2 Pt 1):E183-8.
- 25. Friess E, Wiedemann K, Steiger A, Holsboer F. The hypothalamic-pituitary-adrenocortical system and sleep in man. Adv Neuroimmunol. 1995;5(2):111-25.
- 26. Wiedemann K, Lauer C, Pollmacher T, Holsboer F. Sleep endocrine effects of antigluco- and antimineralocorticoids in healthy males. Am J Physiol. 1994;267(1 Pt 1):E109-14.
- 27. Steiger A, Rupprecht R, Spengler D, Guldner J, Hemmeter U, Rothe B, Damm K, Holsboer F. Functional properties of deoxycorticosterone and spironolactone: Molecular characterization and effects on sleep-endocrine activity. J Psychiatr Res. 1993;27(3):275-84.
- 28. Elenkov IJ, Chrousos GP. Stress hormones, Th1/Th2 patterns, pro/anti-inflammatory cytokines and susceptibility to disease. Trends Endocrinol Metab. 1999;10:359-68.
- 29. Abbas AK, Murphy KM, Sher A. Functional diversity of helper T lymphocytes. Nature. 1996;383(6603):787-93.
- 30. Shanks N, Griffiths J, Anisman H. Central catecholamine alterations induced by stressor exposure: analyses in recombinant inbred strains of mice. Behav Brain Res. 1994;63(1):25-33.
- 31. Webster JI, Tonelli L, Sternberg EM. Neuroendocrine regulation of immunity. Annu Rev Immunol. 2002;20:125-63.
- 32. Dhabhar FS, Miller AH, McEwen BS, Spencer RL. Stress-induced changes in blood leukocyte distribution. Role of adrenal steroid hormones. J Immunol. 1996;157(4):1638-44.
- 33. Moncek F, Kvetnansky R, Jezova D. Differential responses to stress stimuli of Lewis and Fischer rats at the pituitary and adrenocortical level. Endocr Regul. 2001;35(1):35-41.
- 34. Berkenbosch F, van Oers J, del Rey A, Tilders F, Besedovsky H. Corticotropin releasing factor producing neurons in the rat are activated by interleukin-1. Science. 1987;238(4826):524-6.
- 35. Besedovsky H, del Rey A, Sorkin E, Dinarello CA. Immunoregulatory feedback between interleukin-1 and glucocorticoid hormones. Science. 1986;233(4764):652-4.
- 36. Hansen J. Increased breast cancer risk among women who work predominantly at night. Epidemiology. 2001;12(1):74-7.
- 37. Carrillo-Vico A, Guerrero JM, Lardone PJ, Reiter RJ. A review of the multiple actions of melatonin on the immune system. Endocrine. 2005;27(2):189-200.
- 38. Lange T, Perras B, Fehm HL, Born J. Sleep enhances the human antibody response to hepatitis A vaccination. Psychosom Med. 2003;65:831-5.
- 39. Spiegel K, Sheridan JF, Van Cauter E. Effect of sleep deprivation on response to immunization. JAMA. 2002;288(12):1471-2.
- 40. Leproult R, Copinschi G, Buxton O, van Cauter E. Sleep loss results in an elevation of cortisol levels the next evening. Sleep. 1997;20(10):865-70.
- 41. Suchecki D, Lobo LL, Hipolide DC, Tufik S. Increased ACTH and corticosterone secretion induced by different methods of paradoxical sleep deprivation. J Sleep Res. 1998;7(4):276-81.
- 42. Suchecki D, Tiba PA, Tufik S. Hormonal and behavioural responses of paradoxical sleep-deprived rats to the elevated plus maze. J Neuroendocr. 2002;14(7):549-54.
- 43. Drake CL, Roehrs T, Roth T. Insomnia causes, consequences, and therapeutics: an overview. Depress Anxiety. 2003;18(4):163-76.
- 44. Drake C, Richardson G, Roehrs T, Scofield H, Roth T. Vulnerability to stress-related sleep disturbance and hyperarousal. Sleep. 2004;27(2):285-91.
- 45. Cartwright RD, Wood E. Adjustment disorders of sleep: the sleep effect of a major stressful event and its resolution. Psychiatry Res 1991;39(3):199-209.
- 46. Spath-Schwalbe E, Gofferje M, Kern W, Born J, Fehm HL. Sleep disruption alters nocturnal ACTH and cortisol secretory patterns. Biol Psychiatry. 1991;29(6):575-84.
- 47. Perlis ML, Giles DE, Mendelson WB, Bootzin RR, Wyatt JK. Psychophysiological insomnia: the behavioural model and a neurocognitive perspective. J Sleep Res. 1997;6(3):179-88.
- 48. Vgontzas AN, Tsigos C, Bixler EO, Stratakis CA, Zachman K, Kales A, Vela-Bueno A, Chrousos GP. Chronic insomnia and activity of the stress system: a preliminary study. J Psychosom Res. 1998;45(1):21-31.
- 49. Vgontzas AN, Bixler EO, Lin HM, Prolo P, Mastorakos G, Vela-Bueno A, Kales A, Chrousos GP. Chronic insomnia is associated with nyctohemeral activation of the hypothalamic-pituitary-adrenal axis: clinical implications. J Clin Endocrinol Metab. 2001;86(8):3787-94.
- 50. Savard J, Laroche L, Simard S, Ivers H, Morin CM. Chronic insomnia and immune functioning. Psychosom Med. 2003;65(2):211-21.
- 51. Sakami S, Ishikawa T, Kawakami N, Haratani T, Fukui A, Kobayashi F, Fujita O, Araki S, Kawamura N. Coemergence of insomnia and a shift in the Th1/Th2 balance toward Th2 dominance. Neuroimmunomodulation. 2002-2003;10(6):337-43.