Abstracts
Fossil remains of birds belonging to the family Phorusrhacidae were studied in several museums of South America, North America and Europe, the main objective being to characterize this family and solve the chaotic state of the nomenclature and classification of these birds. Reconstruction of some species has been done, with the purpose of having an idea about the size, body weight, posture and habit based in their skeletons. The European species, Ameghinornis minor and Aenigmavis sapea are refuted as belonging to this family. Also several forms described from the Tertiary of Argentina are refuted, because they are based on inadequate segments of the skeleton for a good identification, as is the case of the genera Cunampaia, Smiliornis, Pseudolarus, Lophiornis and Riacama, frequently refered to as belonging to the Phorusrhacidae. The Phorusrhacidae family probably originated in South America, since the end of the Cretaceous, as a result of an endemism formed by the isolation of this landmass. During the end of the Pliocene, with the emersion of the Panama isthmus, the family spread to the North America where at least one species is known Titanis walleri, which perhaps represents the last known species of this family, probably becoming extinct in the beginning of the Pleistocene. A systematic revision has been conducted, dealing with the countless problems of nomenclature, and the Phorusrhacidae is now composed of five subfamilies, which are: Brontornithinae, Phorusrhacinae, Patagornithinae, Psilopterinae and Mesembriornithinae in which 13 genera and 17 species are considered. Characters of all taxa are described and a geochronological distribution of all species is presented.
Phorusrhacidae; Ralliformes; Gruiformes; Tertiary; Giant birds
Foram estudados os fósseis de aves atribuídos à família Phorusrhacidae depositados em diversos museus da América do Sul, da América do Norte e da Europa, com o objetivo principal de caracterizar esta família e reorganizar o estado caótico que até então envolvia a nomenclatura e classificação destas aves. A reconstituição de algumas espécies é feita, com o propósito de formar uma idéia sobre o tamanho, massa corpórea, postura e hábitos com base no esqueleto das mesmas. As formas européias, Ameghinornis minor e Aenigmavis sapea são refutadas como pertencentes à esta família. São refutadas ainda várias formas do Terciário da Argentina, descritas com base em segmentos de esqueleto, insuficientes para uma plena identificação como é o caso dos gêneros Cunampaia, Smiliornis, Pseudolarus, Lophiornis e Riacama, freqüentemente referidos como pertencentes aos Phorusrhacidae. A família Phorusrhacidae certamente originou-se na América do Sul pelo final do Cretáceo, como resultado de um endemismo formado pelo isolamento dessa porção de terra. Pelo final do Plioceno, com a emersão do istmo do Panamá, a família estendeu-se até a América do Norte onde pelo menos uma espécie, Titanis walleri que talvez represente a última conhecida desta família, que extinguiu-se no início do Pleistoceno. A revisão sistemática foi conduzida com inúmeros problemas de nomenclatura e a família Phorusrhacidae passa então a ser constituída de cinco subfamílias, ou seja: Brontornithinae, Phorusrhacinae, Patagornithinae, Psilopterinae e Mesembriornithinae, nas quais se distribuem 13 gêneros e 17 espécies. Os caracteres de todos os táxons são descritos e finalmente é apresentada uma distribuição geocronológica de todas as espécies.
Phorusrhacidae; Ralliformes; Gruiformes; Terciário; Aves gigantes
Systematic revision of the Phorusrhacidae (Aves: Ralliformes)
Herculano M.F. AlvarengaI; Elizabeth HöflingII
IMuseu de História Natural de Taubaté. Rua Colômbia, 99, CEP 12030-520, Taubaté, SP, Brasil. Email: halvarenga@uol.com.br
IIDepartamento de Zoologia, Instituto de Biociências, Universidade de São Paulo, Caixa Postal 11.294, CEP 05422-970, São Paulo, SP, Brazil
ABSTRACT
Fossil remains of birds belonging to the family Phorusrhacidae were studied in several museums of South America, North America and Europe, the main objective being to characterize this family and solve the chaotic state of the nomenclature and classification of these birds. Reconstruction of some species has been done, with the purpose of having an idea about the size, body weight, posture and habit based in their skeletons. The European species, Ameghinornis minor and Aenigmavis sapea are refuted as belonging to this family. Also several forms described from the Tertiary of Argentina are refuted, because they are based on inadequate segments of the skeleton for a good identification, as is the case of the genera Cunampaia, Smiliornis, Pseudolarus, Lophiornis and Riacama, frequently refered to as belonging to the Phorusrhacidae. The Phorusrhacidae family probably originated in South America, since the end of the Cretaceous, as a result of an endemism formed by the isolation of this landmass. During the end of the Pliocene, with the emersion of the Panama isthmus, the family spread to the North America where at least one species is known Titanis walleri, which perhaps represents the last known species of this family, probably becoming extinct in the beginning of the Pleistocene. A systematic revision has been conducted, dealing with the countless problems of nomenclature, and the Phorusrhacidae is now composed of five subfamilies, which are: Brontornithinae, Phorusrhacinae, Patagornithinae, Psilopterinae and Mesembriornithinae in which 13 genera and 17 species are considered. Characters of all taxa are described and a geochronological distribution of all species is presented.
Keywords: Phorusrhacidae, Ralliformes, Gruiformes, Tertiary, Giant birds.
RESUMO
Foram estudados os fósseis de aves atribuídos à família Phorusrhacidae depositados em diversos museus da América do Sul, da América do Norte e da Europa, com o objetivo principal de caracterizar esta família e reorganizar o estado caótico que até então envolvia a nomenclatura e classificação destas aves. A reconstituição de algumas espécies é feita, com o propósito de formar uma idéia sobre o tamanho, massa corpórea, postura e hábitos com base no esqueleto das mesmas. As formas européias, Ameghinornis minor e Aenigmavis sapea são refutadas como pertencentes à esta família. São refutadas ainda várias formas do Terciário da Argentina, descritas com base em segmentos de esqueleto, insuficientes para uma plena identificação como é o caso dos gêneros Cunampaia, Smiliornis, Pseudolarus, Lophiornis e Riacama, freqüentemente referidos como pertencentes aos Phorusrhacidae. A família Phorusrhacidae certamente originou-se na América do Sul pelo final do Cretáceo, como resultado de um endemismo formado pelo isolamento dessa porção de terra. Pelo final do Plioceno, com a emersão do istmo do Panamá, a família estendeu-se até a América do Norte onde pelo menos uma espécie, Titanis walleri que talvez represente a última conhecida desta família, que extinguiu-se no início do Pleistoceno. A revisão sistemática foi conduzida com inúmeros problemas de nomenclatura e a família Phorusrhacidae passa então a ser constituída de cinco subfamílias, ou seja: Brontornithinae, Phorusrhacinae, Patagornithinae, Psilopterinae e Mesembriornithinae, nas quais se distribuem 13 gêneros e 17 espécies. Os caracteres de todos os táxons são descritos e finalmente é apresentada uma distribuição geocronológica de todas as espécies.
Palavras-chave: Phorusrhacidae, Ralliformes, Gruiformes, Terciário, Aves gigantes.
INTRODUCTION
Historicalbackground At the end of the 19th century, Ameghino (1887) described a large, toothless jaw from the Miocene of the Province of Santa Cruz, naming it Phorusrhacos longissimus and assigning it to a new family of toothless mammals. In 1889, Moreno was the first to refer to the giant birds of the Mio-Pliocene from northern Argentina, proposing the term Mesembriornis milneedwardsi for a tibiotarsus, a fibula and a vertebra, associated and incomplete, and Paleociconia australis for an incomplete tarsometatarsus. Two years later, Moreno and Mercerat (1891) recognized for the first time that the mandible described by Ameghino was really that of a bird (they wrongly interpreted it as an upper jaw). In the same year Ameghino (1891a, 1891b) described other birds similar to Phorusrhacos, now recognizing all of them as birds belonging to the group of the Ratitae.
Moreno and Mercerat (1891) described a large amount of fossils of these giant birds from the La Plata Museum, most of which also coming from the Santa Cruz region, as was the case of those described by Ameghino. These authors, however, recognized certain peculiarities of these birds, thus placing them in a new order, the Stereornithes, having some affinities with "los Anseres, de los Herodines y de los Accipitres", dividing this order into four families, which are, the Brontornithidae, Stereornithidae (in which Phorusrhacos is included), Dryornithidae and Darwinornithidae.
The material studied and described by Moreno & Mercerat (1891), at least in part, is made up of segments from skeletons which are of significant diagnostic importance; the work being richly illustrated, thus constituting a landmark for Phorusrhacidae systematics.
A certain rivalry began as to the priority of the terms created by Ameghino (1891a and 1891b), and Moreno & Mercerat (1891). Mercerat himself (1897:227) declares that the "Catálogo de los Pájaros Fósiles de la República Argentina" of which he is co-author, and wherein the presentation done by Moreno is dated the 15th of April, 1891, was first published in mid-May, 1891 (several copies then being distributed in this form), the plates only being terminated on the fifth of August, 1891. Further on he also affirms that Ameghino's publication (1891a), dated as of June the 1st, 1891, was in fact published on the 11th of August, 1891, wherein Ameghino also describes Tolmodus inflatus as a mammal. The second publication of Ameghino (1891b) was printed in December, 1891.
In various subsequent works by various authors (e.g. Andrews, 1896 and 1899; Lambrecht, 1933; Sinclair & Farr, 1932), one observes a strong tendency to give precedence to the terms given by Ameghino over those by Moreno & Mercerat.
Brodkorb (1967), in his revision recognizes the priority of Moreno & Mercerat (1891) over the works of Ameghino of the same year.
In subsequent works, Ameghino (1895 and 1898) describes new fossils of the Phorusrhacidae, creating new terms, often complicating still further the nomenclature of these birds.
Andrews (1896 and 1899), in the detailed study of a certain species, recognizes the close relationship of the Stereornithes with the extant Cariamidae family, discerning a new position, to date still accepted, of the relationship with this family.
Theproposedclassifications One of the first attempts at organizing the classification of these birds was done by Dolgopol de Saez (1927), only with those of the Santacrucian (Mid-Miocene), dividing them into two orders: (1) order Stereornithes with only one family, the Phororhacidae, with two genera, Phororhacos and Psilopterus, and (2) the order Brontornithes, also with one family, the Brontornithidae, with the genera Brontornis, Rostrornis, Liornis and Paleociconia. Dolgopol de Saez (op. cit.), in her classification, gave importance to the format of the ungual phalanges, these being flattened or raised (without specifying on which digit), to the branching or not of the distal foramen of the tarsometatarsus, and to the existence or not of the supratendinal bridge on the tibiotarsus (this without bothering about the frequent losses by fossilization and accidents in collecting).
Patterson & Kraglievich (1960), on studying the forms from the Pliocene, present an important contention on the synonymy, diversity and classification of these birds, placing them in the order Grues Bonaparte, 1857, suborder Cariamae. Moreover, the above mentioned authors propose dividing the Cariamae into two superfamilies, Cariamoidea Stejneger, 1887 and Phororhacoidea Patterson, 1941, and characterize the Phororhacoidea, setting them apart from the Cariamoidea by the following characteristics: (1) a completely desmognate cranium, (2) a relatively high and long rostrum (premaxillar region), (3) ribs without the uncinate processes, (4) very reduced wing bones and loss of the ability to fly, (5) a narrow pelvis with an incomplete pubis and shorter pre-acetabulum portion than the pos-acetabulum one. Further on, Patterson & Kraglievich (op. cit.) divide the Phororhacoidea into the families: Psilopteridae, with the subfamilies Psilopterinae (Psilopterus, Smiliornis and Procariama) and Hermosiornithinae (Hermosiornis), and the family Phororhacidae, with the subfamilies Phororhacinae (Phororhacos, Devincenzia and Onactornis) and Tolmodinae (Tolmodus, Andrewsornis and Andalgalornis). The cited authors (1960:11), moreover, considered the separation of the family Brontornithidae, criticizing, however, the separation of this as an order apart, as proposed by Dolgopol de Saez (1927).
Brodkorb (1967) in his recognized "Catalogue of Fossil Birds", considered the family Phorusrhacidae, giving priority to the spelling Phorusrhacos (Ameghino, 1887) over Phororhacos, this term having been emended by Ameghino (1889) himself, with the subfamilies Brontornithinae (Physornis and Brontornis), Paleociconiinae (Andrewsornis, Palaeociconia and Andalgalornis) and Phorusrhacinae (Phorusrhacos, Onactornis and Titanis), as well as the family Cariamidae with the subfamilies Psilopterinae (Riacama, Smiliornis, Pseudolarus, Psilopterus, Lophiornis and Procariama), Prophororhacinae (Prophororhacos) and Cariaminae (with the extant Cariama and Chunga). The cited author ignored the characteristics by which Patterson & Kraglievich (1960) separated the Cariamoidea from the Phororhacoidea.
In 1981, Mourer-Chauviré described a few bones of birds from the Eocene-Oligocene of France, assigning them to the Ameghinornithinae, a new subfamily of the Phorusrhacidae, the first record of this family for Europe. On complementing her description, the cited author proposes a classification for the suborder Cariamae Fürbringer, 1888, wherein the Phorusrhacidae are divided into six subfamilies, namely, the Brontornithinae (Physornis and Brontornis), Palaeociconiidae (Andrewsornis, Palaeociconia and Andalgalornis), Phorusrhacinae (Phorusrhacos, Onactornis and Titanis), Psilopterinae (Psilopterus, Lophiornis and Procariama), Prophororhacinae (Prophororhacos) and Ameghinornithinae (Ameghinornis). Furthermore, Mourer-Chauviré considers the genera Riacama, Smiliornis and Pseudolarus as insertae sedis.
Geographicalandchronologicaldistribution The greater part of the fossils of the Phorusrhacidae came from Argentina. This does not mean a more southern distribution in South America, but better conditions for the appearance of fossiliferous outcrops in this region, besides the greater technical development in the Paleontology of Argentina, from which one concludes a greater knowledge of fossils in general in that country within South America. Outside Argentina, the Phorusrhacidae birds are known in Uruguay (Kraglievich, 1932; Tambussi et al. 1999), Brazil (Alvarenga, 1982 and 1985a), the Antarctic (Case et al. 1987), and in North America (Brodborb, 1963). More recently, reference is made to these birds in Europe (Mourer-Chauviré, 1981; Peters, 1987), and, possibly still further in the Lower Tertiary, in North America (unpublished material, P. Houde and S. Olson, pers. inf.). These occurrences in the Lower Tertiary in North America and Europe make the biogeographical explanation of the origin and dispersion of the family very difficult.
The oldest fossil record of the Phororhacoidea birds is of Alvarenga (1985), represented by a relatively small-sized form, Paleopsilopterus itaboraiensis from the Mid-Paleocene (Itaboraian) from southeast Brazil, Rio de Janeiro, the Itaboraí Basin. The most recent occurrence is assigned to the limit between the Upper Pliocene and Lower Pleistocene (Late Blancan), from Florida, U.S.A., with a gigantic form, Titanis wagleri, described by Brodkorb (1963).
MATERIAL AND METHODS
The fossilized remains assigned to the Phorusrhacidae and deposited in the collections of the following museums were examined: Museo Argentino de Ciencias Naturales of Buenos Aires, Museo de La Plata, Museu Nacional do Rio de Janeiro, Departamento da Produção Mineral do Rio de Janeiro, Field Museum of Natural History of Chicago, American Museum of Natural History of New York, The Natural History Museum of London, Muséum National d'Histoire Naturelle de Paris, and Forschungsinstitut Senckenberg in Frankfurt.
The material refering to Titanis walleri Brodkorb, 1963, deposited in the Museum of the University of Florida, not only the type, but also several other parts assigned to the species, besides the tarsometatarsus of Devincenzia gallinali Kraglievich, 1932 (type) from the Museu National de Historia Natural of Montevideo, the skull and the bones of the hind-limbs type of Hermosiornis rapax Kraglievich, 1946, deposited in the museum of Mar del Plata, as well as the material whereon the original description of the genus Ameghinornis Mourer-Chauviré, 1981 (part deposited in the museum of Paris and part in the museum of Lyon) was based, were studied from casts kindly ceded by the authorities of these Institutions.
The collection of the Phorusrhacidae compiled by the Princeton University and described by Sinclair & Farr (1932), and recently transfered to the collections of the American Museum of Natural History and of the University of Yale, were studied and compared (those specimens transfered to the University of Yale), based only on the cited authors illustrations.
Amongst the fossils assigned to other groups of South American birds of a medium or large size, the fossils of Argentavis magnificens Campbell Jr. & Tonni, 1980 (Teratornithidae), Opisthodactylus patagonicus Ameghino, 1891, and other rheids, were examined, naturally looking for some mistake in identification. Moreover, the original fossils of Neocathartes grallator (Wetmore, 1944) of the family Bathornithidae (Olson, 1985), besides other original fossils and casts related to this family of the Tertiary from the Northern Hemisphere and very close to the Cariamidae (Cracraft, 1968 and 1971), were examined on a favourable occasion in the National Museum of Natural History, Washington, DC (seeing that the fossils of Neocathartes gallator are not deposited in this museum). Another family of the Lower Tertiary, especially in Europe, and related to the Cariamidae, are the Idiornithidae, which were studied using casts of Elephrocnemus phasianus offered by Cécile Mourer-Chauviré, of the University of Lyon, and from the available illustrations in the publication of this researcher (Mourer-Chauviré, 1983).
The evaluation of the body mass of certain phorusrhacids was done based on the hind-limbs, especially the measurements of the smallest circunference of the diaphysis of the femur and tibiotarsus, which present a direct relationship to the body mass of the bird (Campbell Jr. & Marcus, 1992). In the present work we compared the measurements of some phorusrhacids (diameter of the hind-limb bones) with the same measurements of homologous bones of the ratite birds, or other land birds, with a known mass/size. Several data on bone and body mass measurements of large present-day and fossil birds were obtained in the publications of Amadon (1947) and Wetmore (1967).
Several skeletons of present-day birds were examined as elements for comparison, all from the collection of Museu de Historia Natural of Taubaté (MHNT), amongst which: Struthio camelus, male (MHNT-01) and female (MHNT-1991); Casuarius casuarius, male (MHNT-03) and female (MHNT-1293); Rhea americana, male (MHNT-668); some original bones of the Dinornithidae (MHNT-wt/n.); the cast of a complete skeleton of Aepyornis maximus (MHNT-wt/n.); Cariama cristata, four specimens, two being males (MHNT-1214 and MHNT-1267), one female (MHNT-1136) and the other of undetermined sex (MHNT-78); Opisthocomus hoazin male (MHNT-665).
The anatomical nomenclature used was mainly according to Baumel & Witmer (1993), and in some cases, Howard (1929) and Gilbert et al. (1981).
InstitutionalAbbreviations AMNH, American Museum of Natural History, New York; BMNH, The Natural History Museum, London; DGM, Divisão de Geologia e Mineralogia do Departamento Nacional da Produção Mineral, Rio de Janeiro; FMNH, Field Museum of Natural History, Chicago; MACN, Museo Argentino de Ciencias Naturales Bernardino Rivadavia, Buenos Aires; MGHN, Musée Guimet d'Histoire Naturelle, Lyon; MHNT, Museu de História Natural de Taubaté; MNHN, Muséum National d'Histoire Naturelle, Paris; MLP, Museo de La Plata; MMCN, Museo Municipal de Ciencias Naturales, Lorenzo Scaglia, Mar del Plata; MNHN-M, Museo Nacional de Historia Natural de Montevideo; MNRJ, Museu Nacional do Rio de Janeiro; PUM, Princeton University Museum, New Jersey; SMF, Forschungsinstitut Senckenberg, Frankfurt; TMM, Texas Memorial Museum, Austin; UF, University of Florida, Gainesville.
RESULTS AND DISCUSSION
SizeandbodymasscalculatedforthePhorusrhacidae The Phorusrhacidae are amongst the largest birds that have ever existed on the planet, and Brontornis burmeisteri (Fig. 1A) is, without doubt, the largest phorusrhacid and the largest known bird of the American continent. Its size rivals that of the elephant birds (Aeopyornis maximus) of the Pleistocene-Holocene of Madagascar, classically considered the largest bird that has existed for all time.
The length of the femur, the tibiotarsus and the tarsometatarsus of Brontornis burmeisteri (MLP88, 89 and 91), are very close to the measurements of the corresponding bones of Aepyornis maximus. However, especially the diameters and circuferences of these bones appear to be nearly 10 to 15% smaller in Brontornis.
Dinornisgiganteus, the largest representative of the 13 species of moas (Dinornithidae), known from New Zeeland (Cracraft, 1976), had a body-mass estimated at weighing 230 to 240 kg (Amadon, 1947), or 278 kg (Campbell Jr. & Marcus, 1992), it most certainly having been the tallest bird that existed, apparent by the erect posture and long cervical column. According to the measurements presented by Cracraft (1976), the length of the bones of the hind-limbs of Dinornis giganteus is very near to the length of the same bones in Aepyornis maximus, in the latter, however, the diameter of these bones being much more larger. The body-mass of Aepyornis maximus, according to Amadon (1947), weighed 438 kg, and to Campbell Jr. & Marcus (1992), 542 kg.
Brontornisburmeisteri (Fig. 1A) must have been about 175 cm high at the level of the back, and the head, when well-raised, could have reached around 280 cm high. Its body mass must have weighed approximately 15 to 20% less that of a large specimen of Aepyornis maximus, or in other words 350 to 400 kg. To be more precise in this calculation has the inconvenience of the small number of known specimens of Brontornis burmeisteri, in comparison to the much more numerous fossils of both Aepyornis and Dinornis.
Another bird which was also one of the biggest (in mass) that has ever existed was Dromornis stirtoni, member of an extinct family (Dromornithidae), of giant, cursorial carnivorous birds of the Middle and Upper Tertiary of Australia, which stood nearly as or even higher than Aepyornis maximus (Wroe, 1998). Together with Brontornis burmeisteri, these are the largest birds that have ever existed.
Paraphysornisbrasiliensis, of which the skeleton of the only known specimen having been re-constructed, was calculated at having been around 140 cm high at the back. The head, when well-stretched, reaching 240 cm high. The femurs and tibiotarsi of this bird, when compared to the same bones of a large-sized male specimen of an ostrich (Struthiocamelus) (MHNT1), with a body-mass, when alive, of 130 kg, presented much larger diameters and circumferences, which lead to estimating the weight of Paraphysornisbrasiliensis as having been around 180 kg.
In Phorusrhacos longissimus (Fig. 1C), the diameter of the hind-limb bones was similar to that of the same limbs of a large male ostrich, and so could have had an estimated mass of 130 kg. It was of the same height or maybe even a little higher than Paraphysornis brasiliensis, with a more slender build and possessed notably longer tarsometatarsi.
Patagornismarshi and Andalgalornis steulleti (Fig. 1D), were very near as to size and body-mass, the latter being slightly bigger. The diameter of the leg bones of both was around 15% larger than an adult male of the present-day rhea, Rhea americana, the height of the back being similar, about 90 to 100 cm, but weighing about 45 to 50 kg, seeing that an adult male rhea weighs around 35 kg. The difference in build between a rhea and these phorusrhacids lies in the head being much bigger and heavier in the latter.
The smallest phorusrhacids are to be found in the subfamily Psilopterinae. Psilopterus bachmanni (Fig. 1E), which, when compared with a present-day cariama (Cariama cristata), showed the length of the femur to having been around 40% longer and the tarsometatarsus 40% shorter, the tibiotarsi being approximately of the same length. The diameters and circumferences of these bones are, however, much bigger in Psilopterus.
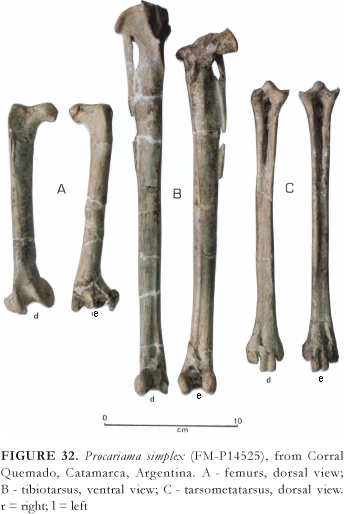
Compared with Cariama cristata, Psilopterus bachmani was of approximately the same height, which was about 60 cm up to the back and 80 cm to the top of the head when well-raised, with a body-mass weighing nearly 5 kg. Psilopterus lemoinei (Fig. 1F), a little bigger, must have had a bulk weighing close to 7 kg. In the same subfamily Psilopterinae, Procariama simplex (Fig. 1G) could have reached nearly 70 cm at the height of the back and have a body-mass weighing near to 10 kg.
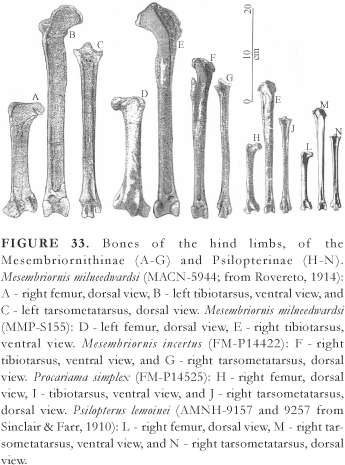
Amongst the Mesembriornithinae, Mesembriornis incertus was built very close to Patagornis marshi and Andalgalornis steulleti, whereas Mesembriornis milneedwardsi (Fig. 1H) was at least 20% bigger and heavier, its bulk reaching almost 70 kg, and the height at the back calculated at 110 or 120 cm, and at the top of the head when well-raised, near to 170 cm.
When attempting to calculate the size and bulk of the birds assigned to the Phorusrhacidae, from Europe, Ameghinornis minor Mourer-Chauviré, 1981, based only on the humerus and coracoid, must have had a build similar to that of Psilopterus lemoinei (a comparison that could be in error through a lack of knowledge regarding the skeleton of the hind-limbs). Aenigmavis sapea Peters, 1987, possessed hind-limb and humerus bones of a size comparable to those of a guan (Penelope obscura), with, however, a much shorter radius and ulna. In this case its body-mass would have been less than 1 kg.
Intra-specificvariations Cracraft (1976) when analizing the skeletons of a large number of kiwis (Apteryx australis) and several species of moas (Dinornithidae), concluded that there is a large degree of intra-specific variation as to the size of the different bones of these birds. Amongst present-day ratites, sexual dimorphism in size is frequent. In Cariama cristata, when observing four skeletons of adult specimens, a difference of 15% in the length of the tarsometatarsus, between the largest and smallest specimens, was noted, with an indication that the largest sizes were from males.
In the case of certain phorusrhacids, such as Brontornis burmeinsteri, a comparison of the tarsometatarsi of two specimens, FMP13259 and MLP91 (lectotype) (Figs. 2C and 2D), both coming from the same geographical region and geological formation, shows them as not to present any anatomical differences, apart from size, wherein the first is around 33% smaller than the second. The idea is that they are examples of intra-specific variation, possibly sexual dimorphism. There is the possibility that they represent two species and a better fossil documentation, however, is necessary to arrive at such a conclusion. It would not be easy to imagine two large predators, extremely similar, disputing the same ecological niche. In birds of prey, dimorphism of such an extent can be observed, as occurs with Harpia harpyja in which adults weigh from 4 kg (generally males) to 9 kg (generally females) (Thiollay, 1994).
Another interesting example occurs with the forms of the genus Psilopterus from the Mid-Miocene (Santa Cruz Formation) of Patagonia. Several species were described based on slight variations in size (Sinclair & Farr, 1932), besides differences as to the height of the upper maxilla and the nasal region. The existance of three species, as these authors believe, is possible, as it would also be possible to accept a single polimorphic species, where the differences represent differences of age and sex, as at present occurs with birds of several groups such as hornbills (Bucerotidae), curassows (Cracidae) and, in a similar way also, the cassowaries (Casuariidae). Another hypothesis is that these shapes might represent temporal differences (even at a specific level), or, in other words, be distributed at different stratigraphic levels.
Shapeandproportionsoftheskeletonandtheirimplicationsonhabits The Phorusrhacidae birds were unable to fly. This conclusion is easily arrived at by examining the proportional size of the wings (Fig. 3) and the body mass, compared with those of present-day forms.
The reduction in wing-size is more pronounced in the larger-sized species. In the diagram depicted in Fig. 3, it can be seen that Paraphysornis, when compared with certain smaller-sized forms, presents a greater reduction of the ulna (Table 1), this reduction appearing to be the main indicator of the loss of the ability to fly. Another example is the comparison of Psilopterus (the smallest Phorusrhacidae) with Cariama cristata, wherein Psilopterus possessed a much larger body-mass, besides an outstanding reduction of the ulna amongst the other wing bones. Moreover, a comparison of the hind-limb bones shows that the shortening of the femur and the relative lengthening of the tarsometatarsus of Cariama in relation to Psilopterus, associated to a smaller bulk appears to be a greater adaptation to running in Cariama. Thus Psilopterus, contrary to the conclusions of Tonni & Tambussi (1988), was unable to fly and was slower than Cariama cristata at running, the latter, on the other hand, achieving long gliding flights of up to some hundreds of meters.
The aptitude for speed in birds when running is proportional to the length of the tarsometatarsus in relation to the tibiotarsus. Thus, in the subfamily Brontornithinae, the largest and heaviest of the Phorusrhacidae, the length of the tarsometatarsus is between 50 to 60% of that of the tibiotarsus, indicating they were slow moving, walking birds, while in Patagornis this proportion is near to 70%, implying much greater agility and speed. Based on this fact, one can deduce that the representatives of the subfamily Brontornithinae must have had necrophagous habits, eating mainly dead or dying animals. This idea can be supported by the information that, in the Tremembé Formation (Taubaté Basin), a lacustrine formation, where the excellent specimen of Paraphysornis brasiliensis came from, periods of drought, with high fish motality, occurred. In the same place, the occurrence of a vulture (Vulturidae) fossil (Alvarenga, 1985b), was also recorded. It is possible that this Phorusrhacidae was hereabouts looking for fishes and other dead animals when, maybe, it sank into the marshy land and succumbed amongst a plentiful supply of fishes and other dead animals.
Another aspect of the skeleton of the Phorusrhacidae is the expressive narrowing of the pelvis, upper maxilla and thorax. From these characteristics we can deduce that these birds often hunted in regions with high vegetation, permitting their greater agility between verticle obstacles. A very narrow upper maxilla would furthermore facilitate the apprehension of small animals hidden amongst trunks or stones.
Another inference derived from the skeleton is from the lacrimal bone (= prefrontal), this forming a large caudal expansion (the supra-orbital process), similar to that which nowadays occurs in hawks (Falconiformes), which certainly affords protection for the eyes against sunrays, thus favoring keeness of sight. From this one supposes that these birds inhabited more or less open sunlit regions and not shaded forests, and certainly hunted by sight.
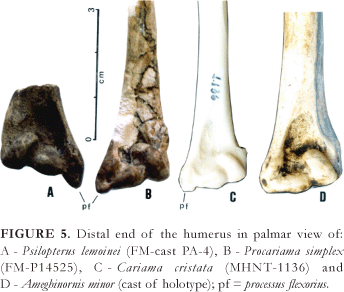
CharacterizationofthePhorusrhacidaefamily Within the order Ralliformes, the family Phorusrhacidae is characterized by: (1) a large or even gigantic build; (2) the shape of the body, especially the premaxila, the pelvis and the thorax being laterally flattened, giving the bird a slim appearance when viewed from the front; (3) a bulky premaxilla, especially high, with a pointed, strong apex, in the form of a hook; (4) a jaw with a very solid symphysis; (5) ample and pervious nostrils (there is no septum); (6) a desmognate palate; (7) the presence of well-developed basipterigoid processes; (8) pterigoids with an evident articulation for the basipterigoid process in the mid-portion; (9) the absence of uncinate processes on the ribs; (10) the pubis atrophied in the cranial half, the same as occurs in hawks (Accipitridae); (11) the reduction of the wings and loss of the ability to fly; (12) a coracoid with the extreme reduction of the procoracoidal and acrocoracoidal processes, possessing an ample scapular facet, in the form of a groove in the apex (Fig. 4B); (13) a humerus with the internal tuberosity bulging proximally, the proximal half of the diaphysis being strongly bent and the processus flexorius distally prominent (Fig. 5); (14) a tarsometatarsus with a triangular-shaped hypotarsus when viewed from below and without tendon grooves; (15) a strongly curved ungual phalanx.
The Cariamidae family, with two extant genera in South America (Cariama and Chunga), and two other very close extinct families that lived in the North American (Bathornithidae) and European (Idiornithidae) MiddleTertiary, are those that present a greater phylogenetic proximity with the Phorusrhacidae (Andrews, 1896; Brodkorb, 1967; Cracraft, 1968 and 1971; Mourer-Chauviré, 1983; Livezey, 1998). They differ, however, through their representatives being of smaller build, the premaxilla not being bulky, the presence of a schizognate palate, the frail mandibular symphysis, the absence of the basipterigoid processes, the relatively wide pelvis, the presence of the uncinate processes on the ribs, the internal tuberosity of the humerus, and a small or non-prominent processus flexorius (Fig. 5).
In the present work, the Ameghinornithinae Mourer-Chauviré, 1981, from Europe, of a size comparable to the smallest Phorusrhacidae, and known only through the coracoid, humerus and carpometacarpus, are excluded from the family Phorusrhacidae. In spite of the reduction of the acrocoracoidal process (Fig. 4A), the coracoid of Ameghinornis presents an excavated scapular facet, this being egg-shaped and extending towards the procoracoidal process, in a shape very typical of the Idiornithidae, thus reminding one especially of Idiornis minor or I. itardiensis (cf. Mourer-Chauviré, 1983: pls. 3 and 4). The humerus of Ameghinornis is similar to that of the Phorusrhacidae as to the prominent ventral tubercle (but of a shape distinct from that of the Phorusrhacidae), and differing in other characteristics such as the distal part of the deltoid crest being much widened and outstanding and the processus flexorius not being prominent distally (Fig. 5D). The carpometacarpus assigned to Ameghinornis reminds one more of that of the Idiornithidae and Cariamidae than that of the Phorusrhacidae, which in turn present a very variable morphology (Fig. 6). Maybe its shape could be interpreted as being primitive for it further resembles that of the Opisthocomidae and Bathornithidae (Olson, 1985:144). The known bones of Ameghinornis compared with Psilopterus (the smallest Phorusrhacidae) and Cariama, are approximately of the same length. The ulna and radius themselves, much more reduced in Psilopterus than in Cariama, appear to be one more characteristic element of the Phorusrhacidae family, by the greater reduction than that of the other wing bones. Unfortunately, the radius and ulna of Ameghinornis are unknown for comparison.
Aenigmavissapea Peters, 1987, another European form described as "Phorusrhacidae" subfamily incertae sedis, and of a much smaller build, is also excluded from the Phorusrhacidae family. Amongst the few osteological characteristics shown by Aenigmavis, the hypotarsus formed by two long and parallel crests differs substancially from that of the Phorusrhacidae. The proportions of the Aenigmavis skeleton are different from those observed in the Phorusrhacidae (Fig. 3): the femur is much longer than the tarsometatarsus, which is only observed in the Phorusrhacidae of the subfamily Brontornithinae (certainly the most derived phorusrhacids). Such a proportion is also observed as common in birds with arboreal habits (e.g. Columbiformes, Psittaciformes, Coraciiformes), as well as in the Galliformes of the Cracidae family, with arboreal habits, such as Pipile and Penelope, thus excluding running habits for Aenigmavis, which must, therefore, remain in insertae sedis. The genera Cunampaia Rusconi, 1946; Smilornis Ameghino, 1891; Pseudolarus Ameghino, 1891; Lophiornis Ameghino, 1891; and Riacama Ameghino, 1899, are also excluded, based on there being few bone segments of very debatable diagnostic value.
Some fossils, still undescribed, coming from the North American Eocene (the Green River Formation), possibly belonging to the Phorusrhacidae (P. Houde, pers. inf.) are also exluded from this family. The examined fossils only consist of wing bones and part of the scapular girdle, of a relatively small-built bird, wherein only the humerus shows some characteristics of the Phorusrhacidae, such as an extremely accentuated bend of the diaphysis, an extremely protruding ventral tubercle, besides a distally prominent processus flexorius. In this humerus, the ventral tubercle is much more developed than in any of the known Phorusrhacidae and is projected more in the anconal than proximal direction. The examined carpometacarpus presents some characteristics common to the Psilopterinae, Cariamidae, Bathornithidae and Opisthocomidae, or, in other words, presents several plesiomorph characteristics, which are of no assistance in an attempt at classification. The observed similarities appear to be more an adaptive convergence due to the loss of the ability to fly, it being extremely premature to classify such birds amongst the Phorusrhacidae.
The Phorusrhacidae thus consists of five subfamilies: the Brontornithinae (Moreno & Mercerat, 1891), Phorusrhacinae (Ameghino, 1889), Patagornithinae (Mercerat, 1897), Psilopterinae (Dolgopol de Saez, 1927) and Mesembriornithinae (Kraglievich, 1932). All the known forms are from South America, except for Titanis walleri, a Phorusrhacinae from North America, which testifies to these birds having emigrated to North America after the elevation of the isthmus of Panama from the Pliocene on.
SYSTEMATICS
The classification herein presented is especially based on the morphology and proportions of the mandibular symphysis and of the tarsometatarsus, for two main reasons: first because one is dealing with resistant bony segments and most often conserved in almost all the species of this family, and also due to the fact that such structures should reflect specialization in food habits as well as differences in the strength and ability of the diverse species.
Order Ralliformes Reichenbach, 1852
Suborder Cariamae Fürbringer, 1888
Family Phorusrhacidae Ameghino, 1889
Phororhacosidae Ameghino, 1889; 1891.
Phororhacidae Lydekker, 1893; Ameghino, 1895.
Phorusrhacidae Brodkorb, 1963,1967; Mourer-Chauviré, 1981.
Brontornithidae Moreno & Mercerat, 1891.
Devincenziidae Kraglievich, 1932.
Stereornithidae Moreno & Mercerat, 1891.
Darwinornithidae Moreno & Mercerat, 1891.
Patagornithidae Mercerat, 1897.
Mesembriorniidae Kraglievich, 1932.
Remarks Ameghino (1887) described Phorusrhacos longissimus based on an incomplete mandible, afterwards (1889) emending the original generic term to Phororhacos, which was widely accepted (Chiappe & Soria, 1990), proposing for this the criation of the family Phororhacosidae. As Brodkorb (1963) warned, the original spelling (Phorusrhacos) has a guaranteed priority. Moreover, Brodkorb emended the name of the family, already previously altered by Lydekker (1893), to the Phorusrhacidae, to which almost all the authors since 1963 have acted in accordance. The terms Phorusrhacos and Phorusrhacidae were definitely made official by the International Commission on Zoological Nomenclature, Opinion 1687: Bulletin of Zoological Nomenclature, 49(2), June, 1992.
Subfamily Brontornithinae Moreno & Mercerat, 1891
Brontornithidae Moreno & Mercerat, 1891.
Brontornithinae; Brodkorb, 1967; Mourer-Chauviré, 1981.
DiagnosisRevised Phorusrhacidae of a gigantic build, reaching over two meters high, heavy and robust. The mandibular symphysis is proportionally shorter, wider and higher than in the other Phorusrhacidae (Fig. 7). The tarsometatarsus is proportionally short, widened and flattened dorso-plantarwise (Fig. 2 and Fig. 8), its length reaching only 50% to 60% of that of the tibiotarsus (Fig. 3). In Brontornis, the dorsomedial extremity of the articular surface of the mesotrochlea of the tarsometatarsus is proximally projected, when viewed in profile (Fig. 8C), whilst in Paraphysornis this portion is more medially expanded. Even though the trochlae of Physornis are as yet unknown, one can assume that the dorsomedial portion of the mesotrochlea in the Brontornithinae may be always expanded and could constitute one more characteristic of the subfamily.
Included Genera Brontornis Moreno & Mercerat, 1891, Physornis Ameghino, 1895, and Paraphysornis Alvarenga, 1993.
Remarks Case et al. (1987) described the fragment of a beak coming from the La Meseta Formation (Upper Eocene ?) from Seymour Island (Antarctic) as a possible portion of the premaxilar region of a Phorusrhacidae. Just by examining the published illustrations, the appearance of the pieces surmises the mandibular symphysis of a Brontornithinae, close to Brontornis. More material and information are required in order to arrive at a precise definition of this possible, as yet unknown, Brontornithinae.
Genus Brontornis Moreno & Mercerat, 1891
Brontornis Moreno & Mercerat, 1891:20,37; Brodkorb, 1967.
Rostrornis Moreno & Mercerat, 1891:20,40; Brodkorb, 1967 (syn. of Brontornis).
Type Species Brontornis burmeisteri Moreno & Mercerat, 1891.
Included Species Only the type species.
Distribution Lower to Mid-Miocene in Argentina.
DiagnosisRevised Certainly the biggest of the Phorusrhacidae, it is the largest bird known from the Americas and one of the largest that has ever existed. The mandibule possesses a proportionally shorter, wider and higher symphysis than Physornis and Paraphysornis (e.g. Alvarenga, 1993: Fig. 1) The internal condyle of the tibiotarsus is medially diverted. The cotyls of the tarsometatarsus are rounded off and the hypotarsus forming a prominent medial crest, having a slightly pronounced lateral edge (e.g. Alvarenga, 1993: Fig. 2). As regards the remaining Brontornithinae, the hypotarsus of Brontornis is placed on a more distal level in relation to the articular cotyles (Fig. 9).
Remarks Liornis Ameghino,1895, with the species L. floweri Ameghino, 1895, and L. minor Dolgopol de Saez, 1927, are evident synonyms of Phorusrhacos Ameghino, 1887; Devincenzia Kraglievich 1932, is surely a robust form of Phorusrhacinae. These two genera were dealt with as synonyms of Brontornis by Brodkorb (1967).
Brontornisburmeisteri Moreno & Mercerat, 1891
Brontornisburmeisteri Moreno & Mercerat, 1891:37; Brodkorb, 1967.
Rostrornisfloweri Moreno & Mercerat, 1891:40; Brodkorb 1967 (syn. of B. burmeisteri).
Brontornisplatyonyx Ameghino, 1895; Brodkorb 1967 (syn. of B. burmeisteri).
Lectotypes The left femur, tibiotarsus, fibula and tarsometatarsus (MLP-88-91), certainly belonging to the same individual, designated by Brodkorb (1967).
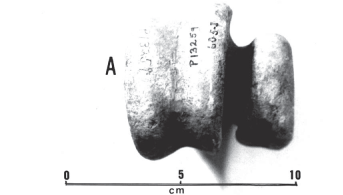
Hypodigm lectotypes; portion of the mandible including the symphysis and part of the right branch (MHNP-1902-6, Fig. 7); two large fragments of the mandibular symphysis (MLP-94-95), mistakenly attributed to the premaxillas by Moreno & Mercerat (1891); quadrate, several complete and incomplete thoracic and caudal vertebras, phalanges and fragments of the hind-limbs (MLP-92-93 and 96117, Fig. 10B); the complete left and the distal end of the right tarsometatarsus (FM-P13259, Fig. 10A); a distal extremity of the right tarsometatarsus (BMNH-A578); distal extremity of the left tarsometatarsus (BMNH-A580); 10 podal and ungual phalanges, the majority belonging to the left foot, apparently from the same specimen (BMNH-A549, Fig. 11); distal end of the left femur (FM-P15309).
HorizonandLocality Lower and Middle Miocene (Santacrucian) of Argentina, Province of Santa Cruz: Lago Argentina, Monte Leon, Monte Observación, Kariaken, La Cueva, Rio Gallegos.
Measurements Table 2.
Illustrations Moreno & Mercerat (1891).
Remarks The trochleae of the right tarsometatarsus of specimen MLP-112 (Fig. 10B), which Moreno & Mercerat (1891) conceived and assigned to Rostrornis floweri, is a mistaken assemblage, wherein the internal trochlea is, in fact, an external left trochlea. Such an error in assemblage also served Dolgopol de Saez (1927) to characterize the genus Rostrornis. An appreciable difference in the size of the tarsometatarsus of specimens FM-P13259 and MLP-91 (lectotype) (Fig. 2C and 2D), shows the second to be around 33% larger than the first, which possibly testifies to a sexual dimorphism, seeing that by the characteristics, both are adults. Two distal fragments of tarsometatarsus from the museum in London (BMNH-A578 and A580) are in accordance with this difference in size (Table 2).
Ameghino (1895) described Brontornis platyonyx, basing it on the much smaller build than B. burmeisteri. His measurements are, however, compatible with the abovementioned variation for the species (Fig. 11).
Genus Physornis Ameghino, 1895
Physornis Ameghino, 1895:576; Brodkorb, 1967.
Aucornis Ameghino, 1898:9; Brodkorb, 1967 (syn. of Physornis).
Type Species Physornis fortis Ameghino, 1895.
Included Species Only the type species.
Distribution Middle and Upper Oligocene of Argentina.
Diagnosis Revised Of a gigantic size, rivaling with Brontornis. A very short and wide mandibular symphysis, characteristically with an almost flat ventral surface in the mid-portion (Fig. 12 and e.g. Alvarenga, 1993: Fig. 1). The lateral cotyl of the tarsometatarsus is almost quadrangular, when viewed proximally (Fig. 13A). The lateral edge of the hipotarsus, when viewed from the rear, forms a prominent crest which distiguishes it well from Brontornis and Paraphysornis (Fig. 13 and e.g. Alvarenga, 1993: Fig. 2).
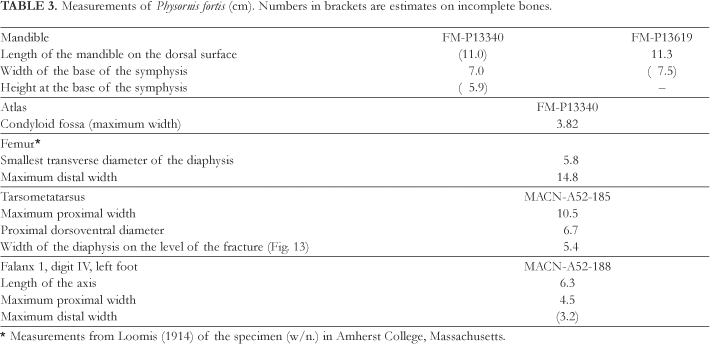
Remarks The genus Physornis and its type species P. fortis, were described by Ameghino in 1895, based on a fragment of the mandible comprehending part of the symphysis and the right branch, this description, unfortunately, not being accompanied by an illustration.
Patterson (1941:52) examined the type of Physornis fortis, nowadays deposited in the museum of London (BMNH-A583), arriving at the conclusion that this is a bony fragment without any morphological characteristic, possibly being the iliac crest of a mammal, thus proposing the rejection of the terms, both for the genus as well as the species, as being indeterminate. We had also the opportunity of examining the aforementioned type material (Fig. 14), arriving at the conclusion that this is undoubtedly a fragment of the symphysis and part of the right branch of a large Phorusrhacidae. The experience acquired in the restoration of the mandible of Paraphysornis (Alvarenga, 1982), contributed to us for the immediate recognition of the texture of a real mandibular symphysis of a Phorusrhacidae. Further, it is evident that Ameghino had prior experience, having examined and described several other mandibular symphysis of Phorusrhacidae, such as, Phorusrhacos longissimus Ameghino, 1887; Phororhacos sehuensis Ameghino, 1891; Phororhacos platygnathus Ameghino, 1891; Tolmodus inflatus Ameghino, 1891, and others, this showing his intimacy with this anatomical portion, which, apparantly, is often conserved in the Phorusrhacidae fossils.
Although it is an almost shapeless fragment and, thus, very inadequate for typifying a species, the type demonstrates, besides the size and the geographical and stratigraphical source, also the flat portion of the ventral surface of the mandibular symphysis, which is characteristic of this genus (Figs. 12A and 14).
Physornisfortis Ameghino, 1895
Physornisfortis Ameghino, 1895:576; Brodkorb, 1967.
Aucorniseuryrhyncus Ameghino, 1898:9; Brodkorb, 1967 (syn. of Physornis fortis).
Type A fragment around 137 cm in length, constituting the ventral portion of the mandibular symphysis and the adjacent part of the right mandibular branch (BMNH-A583: Fig. 14).
Hypodigm Type; a mandibular symphysis, including a part of the left branch (Figs. 12-AC), associated to the caudal portion of the left quadratojugal (Fig. 15A), and to the atlas (Fig. 16A), besides fragments of vertebras, phalanges etc. (FMP-13340). A well conserved madibular symphysis (Fig. 12D), associated to the caudal extremity of the right quadratojugal (FM-P13619). A proximal portion of the left tarsometatarsus (MACN-A-52-185, Figs. 2A and 13). A rostral extremity of the mandibular symphysis (MACN-A-52-186). A phalanx 1 of the toe II from the left foot (MACN-A-52-187); these last three bones were associated and served as type for Ancornis euryrhyncus Ameghino, 1898. A phalanx 1 of the toe IV from the left foot (MACN-A-52-188). In the Museum of Amherst College, Massachussetts, U.S.A., there is also a right femur, incomplete on the proximal extremity, without number (?), refered to and figured by Loomis (1914:226).
HorizonandLocality Middle to Upper Oligocene of Argentina (Deseadan). Province of Santa Cruz, Argentina: Puerto Deseado, Punta Nova, La Flexa, in the levels of occurrence of Pyrotherium. For a better indication of the geological age, see Mac Fadden (1985) and Marshall et al. (1986).
Measurements Table 3.
Illustrations Loomis, 1914: Fig. 149.
Remarks The size of the mandibular symphysis (type: BMNH-A583), originally described by Ameghino (1895), is in perfect agreement with the two measured specimens in the Field Museum of Chicago (Table 3). A flat (not convex) region in the mid-ventral portion of the symphysis is noted in all the specimens.
The branches of the mandible of Physornis, in comparison with those of Phorusrhacos, are really more further apart from the median line, as in the original description by Ameghino (op. cit.), which implies a skull with a wider base. Ameghino perfectly described the only large built Phorusrhacidae of the stratigraphic layers characterized by the presence of Pyrotherium (Deseadan). Thus, there is no reason for the rejection of the genus Physornis and the species P. fortis, as being indeterminate, as suggested by Patterson (1941).
Although the complete shape of the tarsometatarsus of Physornis is unknown, the proximal portion of this is shown to be quite flattened in the dorsoventral extent. Associating this to the short and wide mandibular symphysis, besides the quite large build, it fits in perfectly amongst the Brontornithinae.
Aucornissolidus Ameghino, 1898, considered by Brodkorb (1967) as synonymous of Physornis fortis, was created only based on a proximal extremity of a phalanx (MACN-A-52-110), of a much smaller-sized bird than Physornis fortis, possibly synonymous of Andrewsornis abbotti Patterson, 1941, which should be left as a species inquirenda, seeing that one is dealing with an absolutely insufficient segment to diagnose a species.
The two symphysis examined in the Field Museum of Chicago, showed an interesting difference on the dorsal surface, which, almost flat in FM-P13340, forms a longitudinal channel in FM-P13340 (Figs. 12C and 12D). These differences were interpreted as being individual variation.
Genus Paraphysornis Alvarenga, 1993
Type Species Physornis brasiliensis Alvarenga, 1982.
Included Species Only the type species.
Distribution Upper Oligocene or the Lower Miocene of Southeast Brazil.
Diagnosis Maybe the smallest of the Brontornithinae, the size being comparable to that of a smaller-sized Brontornis burmeisteri (Figs. 1B and 2B). The mandibular symphysis is longer and narrower than in the remaining Brontornithinae (e.g. Alvarenga, 1993: Fig. 1), being, however, proportionally wider and shorter than in the Phorusrhacinae. The cotyles of the tarsometatarsus are slightly quadrangular, especially the inner one (e.g. Alvarenga, 1993: Fig. 2).The lateral edge of the hypotarsus expands until very near to the lateral part of the lateral cotyle, not forming the lateral crest characteristic of Physornis.
Paraphysornisbrasiliensis (Alvarenga, 1982)
Physornisbrasiliensis Alvarenga, 1982.
Holotype The almost complete skeleton of one specimen (Fig. 17), most of the upper maxilla, braincase, pelvis and sternum being missing (DGM-1418-R).
Hypodigm Only the type material.
HorizonandLocality The Tremembé Formation, Taubaté Basin, State of São Paulo, Brazil. Age near to the Upper Deseadean, certainly the Upper Oligocene or the Lower Miocene (Soria & Alvarenga, 1989, and Alvarenga, 1990).
Measurements Tables 1 and 4 ; Alvarenga (1982).
Illustrations Alvarenga (1982, 1993).
Remarks A comparison as to the size of the Brontornithinae, becomes difficult due to the lack of a larger number of individuals and better knowledge of species size variations. The dimension of the tarsometatarsus of Paraphysornis (the only known specimen) is equivalent to that of a small Brontornis (FM-P13259), even though a little less bulky (Figs. 2B and 2C). On the other hand, a comparison of the basal portion of the quadratojugal (Fig. 15), of the atlas (Fig. 16), of the third cervical vertebra and of the proximal portion of the tarsometatarsus of Physornis and Paraphysornis, shows a slight superiority in the size of these in Physornis. However, other specimens, as they come to light, may alter this difference, for the time being established.
Subfamily Phorusrhacinae Ameghino, 1889
Phororhacosidae Ameghino, 1889.
Phororhacidae Ameghino, 1895.
Phororhacinae Kraglievich, 1932.
Phorusrhacinae Brodkorb, 1963; 1967.
DiagnosisRevised Phorusrhacidae, of a gigantic build, having, however, a more slender and lighter constitution, certainly being more nimble, agile and faster (Fig. 1C), than the Brontornithinae. The mandibular symphysis, relatively longer, narrower and not so high (Figs. 18 and 19), is more than twice as long as the width of the base. The tarsometatarsus (Figs. 2E and 2F), relatively long and slender, is always longer than 60% of the length of the tibiotarsus (Figs. 20 and 21).
Included Genera Phorushacos Ameghino, 1887, Devincenzia Kraglievich, 1932 and Titanis Brodkorb, 1963.
Genus Phorusrhacos Ameghino, 1887
Phorusrhacos Ameghino, 1887:24; Bordkorb, 1967.
Phororhacos Ameghino, 1889:659.
Stereornis Moreno & Mercerat, 1891:21, 45; Brodkorb, 1967 (syn. of Phorusrhacos).
Darwinornis Moreno & Mercerat, 1891:26, 60; Brodkorb, 1967 (syn. of Phorusrhacos).
Owenornis Moreno & Mercerat, 1891:25, 64; Brodkorb, 1967 (syn. of Phorusrhacos).
Mesembriornis Moreno, 1889:29; Brodkrob, 1967 (syn. of Phorusrhacos).
Titanornis Mercerat, 1893:5; Brodkorb, 1967 (syn. of Phorusrhacos).
Callornis Ameghino, 1895:574; Brodkorb, 1967 (syn. of Phorusrhacos).
Eucallornis Ameghino, 1901:78; Brodkorb, 1967 (syn. of Phorusrhacos).
Liornis Ameghino, 1895:570; syn. nov.
Type Species Phorusrhacos longissimus Ameghino, 1887.
Included Species Only the type species.
Distribution Lower and Middle Miocene of Argentina.
Diagnosis Of a very big build, being, however, smaller than Devincenzia. The length of the tarsometatarsus is about 70% of that of the tibiotarsus (Figs. 20 and 21), the distal portion of the mid-trochlea not being laterally expanded as is the case of Titanis (Fig. 22).
Remarks Within the subfamily, Phorusrhacos is that which possesses the largest amount of fossils, these coming from the Santacruzian sediments in Argentina. To elaborate a more detailed anatomical diagnosis which would separate Phorusrhacos from both Devincenzia and Titanis becomes, however, difficult through the deficiency in material, seeing that the latter two genera are very badly represented. It is worthwhile pointing out that Devincenzia, with a size which appears to exceed that of Phorusrhacos, comes from more recent levels of Argentina and Uruguay, whereas Titanis, with the same size, is only known from a still more recent terrain of North America.
Phorusrhacoslongissimus Ameghino, 1887
Phorusrhacoslongissimus Ameghino, 1887:24; Brodkorb, 1967.
Phororhacoslongissimus Ameghino, 1889:659.
Stereornisrollieri Moreno & Mercerat, 1891:21, 45; Brodkorb, 1967 (syn. of P. longissimus).
Stereornisgaundryi Moreno & Mercerat, 1891:21,47; Brodkorb, 1967 (syn. of P. longissimus).
Mesembriornisstuderi Moreno & Mercerat, 1891:21, 48; Brodkorb 1967 (syn. of P. longissimus).
Mesembriornisquatrefragesi Moreno & Mercerat, 1891:22, 50; Brodkorb, 1967 (syn. of P. longissimus).
Darwinorniscopei Moreno & Mercerat, 1891:24, 60; Brodkorb, 1967 (syn. of P. longissimus).
Darwinorniszittelli Moreno & Mercerat, 1891:25, 63;
Brodkorb, 1967 (syn. of P. longissimus).
Darwinornissocialis Moreno & Mercerat, 1891:25, 63; Brodkorb, 1967 (syn. of P. longissimus).
Owenornisaffinis Moreno & Mercerat, 1891:25, 64; Brodkorb, 1967 (syn. of P. longissimus).
Owenornislydekkeri Moreno & Mercerat, 1891:25, 64; Brodkorb, 1967 (syn. of P. longissimus).
Phororhacossehuensis Ameghino, 1891:258; Brodkorb, 1967 (syn. of P. longissimus).
Phororhacosplatygnathus Ameghino, 1891:452; Brodkorb, 1967 (syn. of P. longissimus).
Titanornismirabilis Mercerat, 1893:5; Brodkorb, 1967 (syn. of P. longissimus).
Callornisgiganteus Ameghino, 1895:78; Brodkorb, 1967 (syn. of P. longissimus).
Eucallornisgiganteus Ameghino, 1901:78; Brodkorb, 1967 (sin. of P. longissimus).
Liornisfloweri Ameghino, 1895; syn. n.
Liornisminor Dolgopol de Saez, 1927; syn. n.
Type A mandibular symphysis, including part of the right mandibular branch, but lacking the rostral extremity, MLP-118 (originally described as the mandible of a toothless mammal).
Hypodigm Type; several segments of skeletons which served as the type for the various synonyms of the species (MLP-119 to 139, 171 to 182). The species is known from segments of almost all the skeleton, there lacking, however, a better representation of the skull. Ameghino (1895) tells of the observation and measurements of the skull of a specimen in nature, in fragments and encrusted in crumbling rock. The mandible, the rostral extremity of the upper maxilla and a fragment apparently from the caudal portion of the supra-orbital process (BMNH-A529, Figs. 18A, 23B , 23C , 24A and 24C ), coming from this specimen, were collected. Several other segments of the mandible represent the species (Figs. 18 and 19). For P. longissimus there is still lacking an appropriate number of associated bones for a whole reconstruction of the bird. The specimen AMNH-9146 (Fig. 20) represents an almost complete left tarsometatarsus, and the AMNH-9497 (Fig. 21) is represented by some associated bones of a leg.
HorizonandLocality Lower and Middle Miocene (Santacrucian) of Argentina; Santa Cruz Formation, the Province of Santa Cruz: La Cueva, Tagua Quemada, Monte Observación, Rio Shehuen.
Measurements Table 5; Moreno & Mercerat, 1891; Ameghino, 1895.
Illustrations Moreno & Mercerat, 1891 and Ameghino, 1895.
Genus Devincenzia Kraglievich, 1932
Devincenzia Kraglievich, 1932:323, 338.
Onactornis Cabrera, 1939:15; syn. n.
Type Species Devincenzia gallinali Kraglievich, 1932, junior synonym of Devincenzia pozzi (Kraglievich, 1931).
Included Species Only the type species.
Distribution Upper Miocene to the Lower Pliocene of Argentina and Uruguay.
Diagnosis They are the biggest of the known Phorusrhacinae, with a height comparable to that of the largest Brontornithinae, being, however, lighter. The skull is about 65 cm long, equivalent to that of Phorusrhacos, this being possibly flattened dorso-ventrally (Fig. 32A). The tarsometatarsus is substantially sturdier than that of Phorusrhacos (Figs. 2F and 22A).
Remarks The general appearance of the tarsometatarsus of Devincenzia (MNHN-189, Fig. 2F), appears to indicate an intermediate position between the Phorusrhacinae and Brontornithinae, as observed by Kraglievich (1932) himself. However, the appearance of the bony cortex makes it clear that one is dealing with a not yet completely adult individual. As happens with several present-day Ralliformes, especially the Rallidae and Cariamidae, besides other groups of birds, it is normal that the proximal metaphysary portion of the tarsometatarsus reaches, in youngsters, a larger width than that in the adults, associated to a shorter length of the bone. Consequently, one supposes that the tarsometatarsus of Devincenzia, in the adult phase, might be a little longer and with the proximal region slightly narrower than appears in the refered specimen. The origin of this tarsometatarsus (MNHN-189), type of Devincenzia gallinali Kraglievich, 1932, and, consequently, its geological age, are unknown. From the appearance of the bone (and certainly the remains of the encrusted substract), Kraglievich supposed it to have originated from the Arroyo Roman, Rio Negro, Uruguay, and points out the difference of color, especially when compared with the bones coming from Patagonia, without, however, comparing it with specimens from the Huayquerian of north Argentina, studied years before by he himself (1931).
Devincenziapozzi (Kraglievich, 1931). comb. n.
Phororhacospozzi Kraglievich, 1931:306.
Phororhacoslongissimusmendocinus Kragkievich, 1931:314; syn. n.
Devincenziagallinali Kraglievich, 1932:323, 338; syn. n.
Onactornisdepressus Cabrera, 1939:15; Brodkorb, 1967 (syn. of O. pozzi).
Onactornispozzi; Brodkorb,1967:165; syn. n.
Onactornismendocinus; Brodkorb, 1967:166; syn. n.
Type Distal portion of the right tarsometatarsus (Fig. 22A), associated to the ungual phalanx of digit II (MACN-6554 and 6681).
Hypodigm Type; a skull much deformed by dorsoventral flattening, most of the upper maxilla being incomplete (Fig. 23A), together with the proximal phalanges of digits II and III (MLP-37-III-7-8), type of Onactornis depressus Cabrera, 1939; right tarsometatarsus, without the inner trochlea and a large part of the hypotarsus (MNHN-M-189, Fig. 2F), type of Devincenzia gallinali (Kraglievich, 1932); fragment of the madibular symphysis (MACN-6933), described by Kraglievich (1931) as Phororhacos ? aff. platygnathus; proximal portion of a right femur, with the head missing (MACN-6930); distal portion of the left tibiotarsus (MACN-13243).
HorizonandLocality Upper Miocene to the Lower Pliocene of Argentina, Provinces of Buenos Aires and Entre Rios (Huayquerian and "Mesopotamian") and Uruguay (Arroyo Roman ?).
Measurements Table 6; Kraglievich, 1931; Patterson & Kraglievich, 1960.
Ilustrations Kraglievich (1931 and 1932) and Cabrera (1939).
Remarks Tambussi et al. (1999) described an almost complete right tibiotarsus (MNHN-M-1563) about 720 mm long, coming from the Raigon Formation (Pliocene) of south Uruguay. In this specimen the cnemial crests are extremely large and cranially projected, in a similar way to what occurrs in Titanis walleri (UF-7333), and different from what occurs with Brontornithinae. This tibiotarsus should be assigned to the genus Devincenzia and very probably to D. pozzi by the characters of a Phorusrhacinae, the size, age and geographic distribution. Unfortunatelly there is no sufficient material for a definite comparison. Another question would be a better definition of the difference in age between the sediments of the Raigon Formation of Uruguay with the Huayquerian and "Mesopotamian" of Argentina, as well as a new study on the possibility that the tarsometatarsus type of D. gallinali (Kraglievich, 1932), may have also been collected in the Raigon Formation and not in the Arroyo Román as supposed by Kraglievich (1932).
Genus Titanis Brodkorb, 1963
Type Species Titanis walleri Brodkorb, 1963.
Included Species Only the type species.
Distribution Upper Pliocene to Lower Pleistocene of Florida and Texas (U.S.A.).
Diagnosis Size similar to that of Phorusrhacos, with a less sturdy tarsometatarsus than that of Devincenzia. The mid-trochlea, in the more distal portion, is spread out onto the sides, differentiating it from the two aforementioned species (Fig. 22C), this aspect reminding one of the Mesembriornithinae.
Titaniswalleri Brodkorb, 1963
Holotype Distal portion of the right tarsometatarsus (UF-4108).
Hypodigm Holotype; phalanx I of digit III (UF-4109) associated (?) to the holotype; phalanx 1 of digit III (TMM-43060-115); proximal extremity of the left tibiotarsus (UF-7333); complete left carpometacarpus (UF-30003, Fig. 6A); two complete cervical vertebras (C2 and C3); the proximal portion of the right fibula, phalanges 1, 2 and 3 of the left digit III and two phalanges 1 of digit IV (left and right) (UF wt/n.).
HorizonandLocality Upper Pliocene to Lower Pleistocene (end of the Blancanian to the beginning of the Irvingtonian) of Florida, (Inglis, on the border between Gilchrist and Columbia County), and the Pleistocene of Texas (Baskin, 1995).
Measurements Tables 1 and 6, Carr (1981), Chandler (1994) and Baskin (1995).
Illustrations Brodkorb, (1963), Carr (1981), Chandler (1994) and Baskin (1995).
Remarks This is the only Phorusrhacidae known outside South America, one of the most recent species, attesting to these birds crossing over to North America on the forming of the land-bridge connecting North and South America, in the Panama region, at the end of the Pliocene. When compared with other Phorusrhacidae, the examined material indicates a large variation in the size of Titanis, maybe leading one to presume sexual dimorphism.
Subfamily Patagornithinae Mercerat, 1897, revalid. name
Patagornithidae Mercerat, 1897.
Tolmodinae Kraglievich, 1932.
Paleociconiinae Brodkorb, 1967; Mourer-Chauviré, 1981.
DiagnosisRevised A relatively medium-sized Phorusrhacidae, smaller and slimmer than the Phorusrhacinae (Fig. 1D). A long and narrow mandibular symphysis (Figs. 25 and 26). Long and slender tarsometatarsi, more than 70% of the length of the tibiotarsus. The tibiotarsi and tarsometatarsi are proportionally more slender than those of the Phorusrhacinae.
Included Genera Patagornis Moreno & Mercerat, 1891, Andrewsornis Patterson, 1941, and Andalgalornis Patterson & Kraglievich, 1960.
Remarks Brodkorb (1967) considered the term Paleociconia revalidated by Moreno & Mercerat (1891), and based on this created the subfamily Paleociconiinae. The genus Paleociconia Moreno, 1889, having as type-species Paleociconia australis assigned by Richmond (1901), is based on a distal half of a left tarsometatarsus, once again described and also pictured by Moreno & Mercerat (1891). Patterson & Kraglievich (1960:68), comment on the extremely superficial diagnosis, without any illustration, by Moreno (1889), and consider Paleociconia australis as nomen nudum. Kraglievich (1931:305) and Brodkorb (1967), consider the genus Paleociconia revalidated by Moreno & Mercerat (1891), having as type-species Paleociconia cristata Moreno & Mercerat, designated by Rovereto (1914:163). It so happens that Paleociconia cristata Moreno & Mercerat, 1891, is based on the very resumed description of two broken cervical vertebras, of a much smaller species, possibly a Psilopterinae, without the characteristics to permit a precise identification and so should be considered as species inquirenda. One ungual phalanx ilustrated by Moreno & Mercerat (PL. XIX12) is only assigned to Paleociconia cristata, without any description, thus being also insufficient to validate the taxon. The term Paleociconia becomes even more confused when Lydekker (1891) committed a grave mistake while describing and picturing the distal extremity of a tarsometatarsus of a true Ciconiidae of the Pleistocene of Lagoa Santa, Brazil, on assigning it to Paleociconia australis Moreno, 1889. In spite of the commentaries emitted by the abovementioned authors, Moreno (1889) had created the genera Mesembriornis and Paleociconia, and nowadays, knowing the respective material on which these genera were based, it is known that they are synonyms and that Paleociconia (Moreno, 1889:29) is a junior synonym of Mesembriornis (Moreno, 1889:30), which in turn is an available name, with type material also available for new studies, and so cannot be rejected as nomen nudum.
Genus Patagornis Moreno & Mercerat, 1891 revalid. name
Patagornis Moreno & Mercerat, 1891 (May).
Tolmodus Ameghino, 1891 (June); Brodkorb, 1967 (syn. of Paleociconia).
Morenomerceraria Lambrecht, 1933; Brodkorb, 1967 (syn. of Paleociconia).
Paleociconia Brodkorb, 1967; Mourer-Chauviré, 1981.
Type Species Patagornis marshi, Moreno & Mercerat, 1891. Designated by Richmond, 1902.
Included Species Only the type species.
Distribution The Lower and Middle Miocene of Argentina.
Diagnosis The antorbital fenestra with the rostral edge moderately inclined (much inclined in Andrewsornis and little inclined in Andalgalornis) (Figs. 26A, 26B and 26C). The dorsal portion of the nostrils is very outstanding. Slightly curved mandibular symphysis (the apex is not raised) (Fig. 26B).
Remarks Patagornis Moreno & Mercerat, 1891 (May-August) has priority over Tolmodus Ameghino, 1891 (June), as held by Mercerat (1897:227) on the question.
Patagornismarshi Moreno & Mercerat, 1891 revalid. name
Patagornismarshi Moreno & Mercerat, 1891 (May).
Tolmodusinflatus Ameghino, 1891 (June); Brodkorb, 1967 (syn. of Paleociconia cristata).
Phororhacosinflatus; Ameghino, 1891 (August); Andrews, 1899; Brodkorb, 1967 (syn. of Paleociconiacristata).
Paleociconiacristata; Brodkorb, 1967.
Lectotype (by present designation): A mandibular symphysis lacking the rostral extremity (MLP-143), described and portrayed by Moreno & Mercerat as a premaxilla.
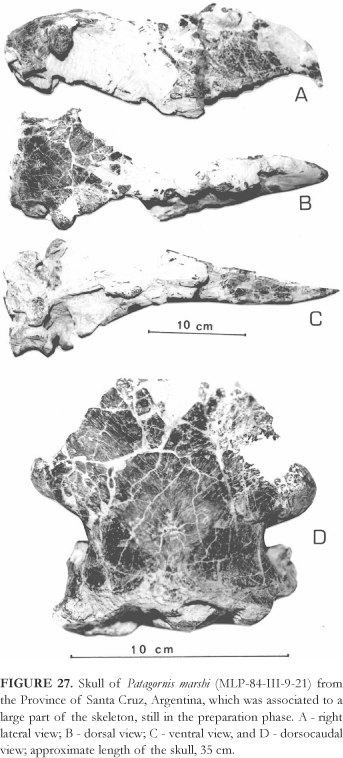
Hypodigm One of the best represented Phorusrhacidae by the amount of fossils. Together with the lectotype, several segments of the skeleton, amongst which various bone segments from the hind-limbs, are described and well portrayed by Moreno & Mercerat (1891), of which at least the majority could belong to the same individual. (MLP-144 to 158). An almost complete and well-preserved skeleton (BMNH-A516), is described in details by Andrews (1899). Another almost complete, although badly deformed skeleton is at the Museo de La Plata (MLP-84-III-9-21), and includes the skull (Fig. 27). A parcial skeleton (AMNH-9264) was well-portrayed by Sinclair & Farr (1910: PL. XXXII), although the authors did not identify it. A complete and isolated tarsometatarsus (FM-P13213) was the only long bone of a young Phorusrhacidae examined in the present work, the proximal epiphysis being completely unfused to the diaphysis, and the total length (243 mm) almost 90% of that of an adult. A fragmment of the rostral portion of the mandibular symphysis of a Phorusrhacidae (BMNH-586) is described and portrayed by Ameghino (1891b) as belonging to the extremity of the premaxilla of Opisthodactilus patagonicus, when in fact it should be assigned to a mandibular symphysis of Patagornis marshi.
HorizonandLocality Lower to Middle Miocene (Santacrucian) of Argentina, Province of Santa Cruz, the Santa Cruz Formation: Monte Observación, Tagua Quemada, La Cueva.
Measurements Tables 1 and 7 ; Andrews (1899).
Illustrations Moreno & Mercerat (1891: PL. XIV and XV), Andrews (1899), Ameghino (1899), and Sinclair & Farr (1932: PL. XXXII).
Genus Andrewsornis Patterson, 1941
Type Species Andrewsornis abbotti Patterson, 1941.
Included Species Only the type species.
Distribution Middle to Upper Oligocene of Argentina.
DiagnosisRevised The largest of the known Patagornithinae. On the skull, the rostral edge of the antorbital fenestra is very inclined. When compared with Patagornis and Andalgalornis, the mandibular symphysis is proportionally smaller and the mandibular fenestra bigger and oval-shaped (Fig. 26A).
Andrewsornisabbotti Patterson, 1941
Type Incomplete skull, mandible (Fig. 26A), omal extremity of a coracoid, the second and ungual phalanges of digit II (FM-P13417).
Hypodigm Type; a mandibular symphysis (FM-P13383) (Figs. 25A, 25B and 25C); a left femur with only the internal condyle missing (FM-P14678) (Figs. 25D, 25E and 25F).
HorizonandLocality Middle and Upper Oligocene of Argentina, (Deseadan); Cabeca Blanca (Chubut), Pico Truncado (Santa Cruz).
Measurements Table 7 ; Patterson, (1941).
Illustrations Patterson (1941).
Remarks The mandibular symphysis of specimen FM-P13383 (Figs. 25A, 25B and 25C), is smaller than that of the type specimen and the femur (FM-P14678) (Figs. 25D, 25E and 25F) is still smaller than the corresponding bone of Patagornis marshi. However, the measurements of the type specimen lead one to believe it to have been a larger bird. This is certainly one more example of intraspecific variation in these birds, and possibly even sexual dimorphism. A phalanx of digit II (MACN-A-52-110), whereon Ameghino (1898) described Aucornis solidus, is very probably attributed to this species and the name of which naturally could have priority. However, the diagnostic value of this phalanx is very debatable, as also one can question the correct geographical and stratigraphical origin of the specimen as well. It is thus preferable in this case to consider Aucornis solidus Ameghino, 1898 as a species inquirenda.
Genus Andalgalornis Patterson & Kraglievich
Type Species Andalgalornis ferox Patterson & Kraglievich 1960 (junior synonym of A. steulleti Kraglievich 1931).
Included Species Only the type species.
Distribution Upper Miocene and Lower Pliocene of Argentina.
DiagnosisRevised Upper maxilla proportionally higher than in the rest of the Patagornithinae, with the dorsal portion of the nostrils not raised. The rostral edge of the antorbital fenestra is almost straight and verticalized. The mandibular symphysis is longer, with the apex curved dorsally (Fig. 26C). The cervical vertebrae and especially the cervicodorsal ones, are sturdier than in Patagornis, and comparing herewith to the pelvis, the iliac crests are dorsally joined, forming a dorsal projection which is more prominent in the region above the acetabulum (Fig. 28B).
Andalgalornissteulleti (Kraglievich, 1931)
Phororhacossteulleti Kraglievich, 1931:312.
Phororhacosdeautieri Kraglievich, 1931:312; Brodkorb, 1967 (syn. of Andalgalornis steulleti).
Andalgalornisferox Patterson & Kraglievich, 1960; syn. n.
Andalgalornissteulleti; Brodkorb, 1967:162.
Type phalanx 1 of digit IV, left side (MACN-4244).
Hypodigm Type; distal portion of the right femur (MACN-6932), type of Phororhacos deautieri Kraglievich, 1931. Partial skeleton consisting of the skull, mandible, pre-synsacral vertebrae (except the atlas), the pelvis and some broken ribs (FM-P 14357), type of Andalgalornis ferox Patterson & Kraglievich, 1960.
Horizonandsites Upper Miocene to Lower Pliocene of Argentina: Andalgala Formation, "Mesopotamian" (= Huayquerian ?), the Entre Rios and Catamarca (Chiquimil) Provinces.
Measurements Table 7 , Patterson & Kraglievich (1960).
Illustrations Kraglievich (1931), Patterson & Kraglievich (1960).
Remarks The fossils of Phororhacos steulleti and P. deautieri come from the "Mesopotamian", whilst those of Andalgalornis ferox from Huayquerian (the Andalgala Formation), possibly different ages of the Upper Miocene and (or) Lower Pliocene (Tonni, 1980); Noriega (1995) believes that the "Mesopotamian" is within the Huayquerian. It is possible that the synonymous species herein became distinct (in time), but there is not enough material to corroborate such a hypothesis.
Subfamily Psilopterinae Dolgopol de Saez, 1927
Pelecyornidae Ameghino, 1891.
Psilopteridae Dolgopol de Saez, 1927.
Psilopteriidae Kraglievich, 1932.
Psilopterinae Patterson & Kraglievich, 1960; Brodkorb, 1967; Mourer-Chauviré, 1981.
DiagnosisRevised A relatively small-built Phorusrhacidae, with thinner legs, and a relatively light and graceful body. Relatively big nostril openings; the rostral edge of the antorbital fenestra is well slanted. The mandible possesses a much lengthened fenestra, sometimes divided forming two fenestras. The mandibular symphysis is relatively small when compared to the total length of the mandible. The cervical vertebras are proportionally longer and narrower than in the remaining subfamilies. The length of the tarsometatarsus is about 70 to 75% of that of the tibiotarsus. The leg bones are proportionally slimmer than in the Patagornithinae and the other Phorusrhacidae.
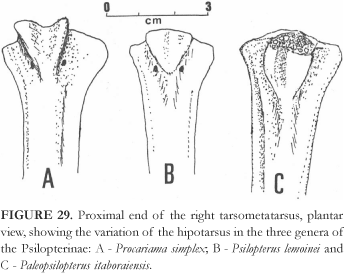
Included Genera Psilopterus Moreno & Mercerat, 1891, Procariama Rovereto, 1914 and Paleopsilopterus Alvarenga, 1985.
Genus Psilopterus Moreno & Mercerat, 1891
Psilopterus Moreno & Mercerat, 1891 (May-August).
Pelecyornis Ameghino, 1891 (December); Brodkorb, 1967 (syn. of Psilopterus).
Staphylornis Mercerat, 1897; Brodkorb, 1967 (syn. of Psilopterus).
Type Species Psilopterus comunis Moreno & Mercerat, 1891, designated by Richmond, 1902 (a junior synonym of Psilopterus bachmani Moreno & Mercerat, 1891).
Included Species P. bachmani (Moreno & Mercerat, 1891), P. lemoinei (Moreno & Mercerat, 1891), P. affinis (Ameghino, 1899) and P. colzecus (Tonni & Tambussi, 1988).
DiagnosisRevised The smallest Psilopterinae. A rather high intercotylar tubercle on the tarsometatarsus, and the lateral edges of the hypotarsus are little or not spread (Fig. 29B).
Distribution Mid-Oligocene to Upper Miocene of Argentina.
Psilopterusbachmanni (Moreno & Mercerat, 1891) comb. n.
Patagornisbachmanni Moreno & Mercerat, 1891 (pp. 24 and 58).
Psilopteruscommunis Moreno & Mercerat, 1891 (pp. 26 and 68); Brodkorb, 1967.
Psilopterusintermedius Moreno & Mercerat, 1891 (pp. 26 and 68); Brodkorb, 1967 (syn. of P. communis).
Phororhacosdelicatus Amegino, 1891; Brodkorb, 1967, (syn. of P. communis).
Pelecyornispueyrredonensis Sinclair & Farr, 1932; Brodkorb 1967 (syn. of P. communis).
Lectotype (by present designation): proximal portion of the left tarsometatarsus (MLP-168).
Hypodigm lectotype. A proximal and a distal portion of a femur (MLP-165-166) and a distal portion of the tibiotarsus, all of the left side (MLP-167), which appear to be associated to the lectotype. An almost complete skeleton (PUM-15.904), described and pictured by Sinclair & Farr (1932).
HorizonandLocality Middle Miocene (Santacrucian) of Argentina, Santa Cruz Province: Santa Cruz, Lake Pueyrredon, Monte Observación, La Cueva.
DiagnosisRevised Perhaps the smallest of all the known Phorusrhacidae (Fig. 1E), estimated at being between 70 to 80 centimeters high, rivalling only Psilopterus affinis in size. Relatively low skull and upper maxilla, in height (Fig. 26I), presenting a certain similarity with the Mesembriornithinae. Differences with P. lemoinei as to the different segments of the skeleton are well described by Sinclair & Farr (1932). A peculiarity of P. bachmanni is to be found in the extremely slanted rostral edge of the antorbital fenestra, very different from P. lemoinei (Figs. 26G and 26I).
Measurements Table 8 and Sinclair & Farr (1932).
Ilustrations Sinclair & Farr (1932) and Moreno & Mercerat (1891).
Remarks Patagornis bachmanni has priority over Psilopterus communis (both of Moreno & Mercerat, 1891). Psilopterus minutus Ameghino, 1981, dealt with as a separate species by Brodkorb (11967), is represented only by an incomplete tarsometatarsus, inseparable from P. bachmanni (v. Sinclair & Farr, 1932:188).
Psilopteruslemoinei (Moreno & Mercerat, 1891) comb. n.
Patagornislemoinei Moreno & Mercerat, 1891 (pp. 23 and 58).
Psilopterusaustralis Moreno & Mercerat, 1891 (pp. 26 and 68); Brodkorb, 1967.
Pelecyornistubulatus Ameghino, 1895; Brodkorb, 1967 (syn. of Psilopterus australis).
Phororhacosmodicus Ameghino, 1895; syn.n.
Staphylornisgallardoi Mercerat, 1897; Brodkorb, 1967 (syn.? of Psilopterus australis).
Staphylorniserythacus Mercerat, 1897; Brodkorb, 1967 (syn.? of Psilopterus australis).
Pelecyornistenuirostris Sinclair & Farr, 1932; Brodkorb, 1967 (syn. of Psilopterus australis).
Lectotype (by present designation): Distal end of a right tibiotarsus (MLP-162).
Hypodigm Besides the lectotype and the material that served as type for the diverse synonyms, a plentiful and splendid material is discribed and pictured by Sinclair & Farr (1932). An ungual phalanx (MLP-164), described and pictured by Moreno & Mercerat (1891) as belonging to this species, certainly belongs to Patagornis marshi.
HorizonandLocality Middle Miocene (Santacrucian) of Argentina, Santa Cruz Province: Santa Cruz, Killik Aike, Monte Observación, Take Harvey, La Cueva, Corriguen Kaik, Tagua Quemada.
DiagnosisRevised Larger-sized than P. bachmanni (Fig. 1F), with the skull and upper maxilla notably higher (Figs. 26F, 26H and 26I), and a less slanted rostral edge of the antorbital fenestra. Other differences in the diverse segments of the skeleton are described and pictured by Sinclair & Farr (1932).
Measurements Table 8 and Sinclair & Farr (1932).
Illustrations Sinclair & Farr (1932).
Remarks One of the best represented Phorusrhacidae by the relative abundance of fossils. Some observed discrepancies, especially in the skulls (Figs. 26F, 26G, 26H and 26I), should be attributed to differences in age or sex. Psilopterus lemoinei and P. bachmanni are herein dealt with as being two very similar species that appear to have lived together, disputing very similar niches. The most important difference between the two is in the size and proportions of the upper maxilla, and the differences in slant of the rostral edge of the antorbital fenestra. A deeper study might lead to different conclusions, as to the number of species of the genus Psilopterus from the Santacruzian of Argentina, as was discussed before under the sub-title: "intra-specific variations".
Psilopterusaffinis (Ameghino, 1899) comb. n.
Phororhacosaffinis Ameghino, 1899.
Type The right tarsometatarsus lacking a segment of diaphysis (MACN-A-52-184).
Hypodigm Only the type material.
HorizonandLocality Guaranitica Formation of Patagonia (= Deseado Formation). The Middle to Upper Oligocene of Argentina (Deseadan), Chubut Province: Cabeca Branca.
Measurements Table 8.
Diagnosis Small size, rivalling Psilopterus bachmanni. It differs from all the others of the same kind as regards the upper part of the hipotarsus, where a transversal groove separates this from the cotylar surface.
Remarks Treated as synonymous of Andrewsornis abbotti by Brodkorb (1967), just because it came from the same stratigraphic levels, it differs from this latter by being of a much smaller build. Brodkorb (op. cit.), furthermore, cites the epithet affinis as being pre-occupied by Owenornis affinis Moreno & Mercerat, 1891, a junior synonymon of Phorusrhacos longissimus, an unfounded concept, seeing that they are perfectly distinct genera. Without doubt, this is the least known species amongst the Phorusrhacidae. It is possible that, with new material and better study, the meagre and insufficient material based whereon Ameghino (1899) decribed other species coming from the "Guaranítica" (Deseadan) Formation of Patagonia, could be clarified: Riacama caliginea, Smiliornis penetrans and Pseudolarus guaraniticus. Some of these could possibly be synonymous with Psilopterus affinis, whereupon to the latter may be added more peculiarites which would justify separating it into a genus apart.
Psilopteruscolzecus Tonni & Tambussi, 1988
Holotype The incomplete skeleton of an individual (MLP-76-VI-12-2) consisting of the right incomplete mandibular symphysis, humerus, radius and ulnar, the right femur, the diaphysal portion of the tibiotarsus and tarsometatarsus, and the distal portion of the left tarsometatarsus together with the repective phalanges.
Hypodigm Only the type material.
HorizonandSite The Upper Miocene (Chasicoan) of Argentina: Arroyo Chasicó Formation, Province of Buenos Aires; Partido de Villarino.
Diagnosis A size similar to that of P. lemoinei, representing the most recently known form of the genus. The most outstanding differences are in details of the trochlea of the tarsometatarsus with the other congenerous forms, are pointed out by Tonni & Tambussi (1988).
Measurements Table 8 and Tonni & Tambussi (1988).
Illustrations Tonni & Tambussi (1988).
Genus Procariama Rovereto, 1914
Type Species Procariama simplex Rovereto, 1914.
Included Species Only the type species.
Distribution Upper Miocene to Lower Pliocene of Argentina. (Catamarca Province).
DiagnosisRevised It is very similar to Psilopterus, being distiguished from this by the larger size and sturdier build (Fig. 1G), and by a slight difference in the proportion of the bones of the hind-limbs, where the femur is proportionally shorter (Patterson & Kraglievich, 1960:16). The wing bones of Procariama are also proportionally smaller (Fig. 3). In the tarsometatarsus, the hipotarsus forms two expansions (one lateral the other medial) in the most proximal part, this thus looking like two crests (Fig. 29A), different from both Psilopterus and Paleopsilopterus.
Procariamasimplex Rovereto, 1914
Lectotype Incomplete skeleton constituted by an incomplete skull, pelvis, proximal and distal portions of the left femur, distal portion of the right tibiotarsus, proximal and distal portions of the right tarsometatarsus, foot and ungual phalanges of the left foot (almost complete) and fragments of the phalanges of the right foot (MACN-8225), designated by Patterson & Kraglievich (1960).
Hypodigm Lectotypes; right femur distally incomplete, distal portion of the left tarsometatarsus and some associated phalanges (MACN-6939); an almost complete skeleton in an excellent state of conservation (FM-P 14525) (Figs. 30, 31 and 32).
HorizonandLocality Upper Miocene and Lower Pliocene (Huayquerian) of Argentina: Catamarca Province: Andalgala, Corral Quemado (Belem), Chiquimil, Rio Santa Maria (Estratos Araucanos).
Measurements Table 8 and Patterson & Kraglievich (1960).
Illustrations Rovereto (1914).
Remarks The similar size to that of Mesembriornis incertus (Figs. 33F and 33G), plus the same stratigraphic occurrence, could lead to confusion amongst the fossils of these species, which, in fact, occurred with Rovereto (1914) in his original description. See the remarks related to Mesembriornis incertus.
Genus Paleopsilopterus Alvarenga, 1985
Type Species Paleopsilopterus itaboraiensis Alvarenga, 1985.
Included Species Only the type species.
Distribution Middle Paleocene of Brazil (Rio de Janeiro).
Diagnosis Heavier built than Psilopterus, similar to Procariama. The intercotylar tubercle of the tarsometatarsus is wide and rounded, and lower than in the abovementioned genera. The hypotarsus is laterally and distally expanded, covering the proximal foramina when viewed from below (Fig. 29C).
Paleopsilopterusitaboraiensis Alvarenga, 1985
Type Proximal portion of the right tarsometatarsus (MNRJ-4040-V).
Hypodigm Type; left and right tibiotarsus, proximally deformed and incomplete (Paratypes, DGM-1431-R).
HorizonandLocality Middle Paleocene (Itaboraian) of southeastern Brazil: Rio de Janeiro, Itaboraí.
Measurements Table 8 and Alvarenga (1985 a).
Illustrations Alvarenga (1985 a).
Remarks It is the oldest Phorusrhacidae known.
Subfamily Mesembriornithinae Kraglievich, 1932 stat. n.
Mesembriorniidae Kraglievich, 1932.
Hermosiornidae Rovereto, 1914; Brodkorb, 1967 (syn. of Prophorhacinae).
Hermosiorniidae Kraglievich, 1932
Hermosiornithidae Wetmore, 1934.
Hermosiornithinae Patterson & Kraglievich, 1960.
Prophororhacinae, Brodkorb, 1967; Mourer-Chauviré, 1981.
DiagnosisRevised This is a relatively middle-sized Phorusrhacidae. The upper maxilla is relatively low, especially in the middle part, being rostrally lengthened (Fig. 1H and Fig. 26D). There is a short and relatively low mandibular symphysis. The coracoid is anchylosed to the clavicle (Figs. 4G and 4H). On the tibiotarsus, the apex of the internal condyle is more prominent and proximally bent, thus forming a more acute angle (when seen in internal view), with the diaphysis. They are the slimmest of the Phorusrhacidae, in which the tarsometatarsus reaches around 80 to 85% of the length of the tibiotarsus (Fig. 33), and wherein the middle trochlea is distally spread, with a width equal or more than the smallest transverse diameter of the diaphysis. The Mesembriornithinae are amongst the most recent of the Phorusrhacidae (Fig. 34).
Included Genera Mesembriornis Moreno, 1889 (unique).
Genus Mesembriornis Moreno, 1889; revalid. name.
Mesembriornis Moreno, 1889:29
Paleociconia Moreno, 1889:30.
Prophororhacus Rovereto, 1914:114.
Hermosiornis Rovereto, 1914:163.
Type Species Mesembriornis milneedwardsi Moreno, 1889, designated by Richmond, 1902.
Included Species The type species and M. incertus Rovereto, 1914.
Distribution Upper Miocene to Upper Pliocene of Argentina.
Diagnosis The same as for the subfamily.
Mesembriornismilneedwardsi Moreno, 1889 revalid.name
Mesembriornismilneedwardsi Moreno, 1889:29; Moreno & Mercerat, 1891; Brodkorb, 1967 (syn. of Prophororhacos australis).
Paleociconiaaustralis Moreno, 1889:30; Moreno & Mercerat, 1891.
Driornispampeanus Moreno & Mercerat, 1891 (part: only femur).
Hermosiornismilneedwardsi Rovereto, 1914.
Hermosiornisrapax Kraglievich, 1946; syn. n.
Prophororhacosaustralis Brodkorb, 1967:172.
Type The centrum of a cervical vertebra associated to the proximal part of both the right tibiotarsus and fibula (MLP-140-142).
Hypodigm Type; distal half of the left tarsometatarsus, lacking the external trochlea (MLP-87 type of Paleociconia australis Moreno, 1889); an almost complete skeleton, lacking especially the skull (MACN-5944); almost complete skeleton, including the skull and mandible, besides the left and incomplete humerus, radius and ulna, the femurs, and the right tibiotarsus and tarsometatarsus (MMP-S155); distal fragment of the right femur (MLP-170).
HorizonandLocality The Middle and Upper Pliocene of Argentina (Montehermosan), Province of Buenos Aires; the Monte Hermoso and Chapadmalal Formations; Monte Hermoso, Rio Loberia.
Measurements Table 9, Rovereto (1914), Kraglievich (1946) and Patterson & Kraglievich (1960).
Illustrations Rovereto (1914) and Kraglievich (1932: PL. LVIII and 1946).
Remarks Only a slighter larger size of specimen MMP-S155 does not justify the Kraglievich's (1946) specific separation of Hermosiornis rapax. Patterson & Kraglievich (1960:19) commented that specimens MACN-5944 and MLP-140-142 may belong to the same individual, which seems to be very probable. The skull of specimen MMP-S155 (Fig. 26D) is the only known one of Mesembriornithinae and even though pictured in Kraglievich's (1946), without comments, it is on some parts reconstructed; the rostral extremity, as well as the nostril region, are reconstructions; the height of the upper maxilla, in the middle part, is well defined but the length seems to be slightly increased in the reconstruction, mainly when considering the conserved mandible; the appearance of this skull seems to be more similar to that of Psilopterus bachmanni (Fig. 26I) than that represented in Kraglievich's (1946:109) restoration. (Julia Clark, pers. inf., 1999).
Mesembriornisincertus (Rovereto, 1914) comb. n.
Prophororhacosincertus Rovereto, 1914; Brodkorb, 1967.
Type A dorsal vertebra; a fragment of the omal portion of the left coracoid; each the right, humerus, ulna and part of the radius; distal part of the right tarsometatarsus and phalanges 1 of digits III and IV and phalanx 3 of digit II (MACN-6934).
Hypodigm Type; the fragment of a cervical vertebra (MACN-6931); a tibiotarsus associated to the fibula, tarsometatarsus (Fig. 33F and 33G) and complete digits I and II, all from the right side (FM-P14422); a left tarsometatarsus, with the proximal portion missing, and phalanges 1 and 2 of digit II (MACN-6737).
HorizonandLocality The Upper Miocene to the Lower Pliocene, of Argentina (Huayquerian): Catamarca Province (Andalgala, Corral Quemado).
Diagnosis Much smaller than M. milneedwardsi.
Measurements Table 9.
Illustrations Rovereto (1914).
Remarks As commented by Patterson & Kraglievivh (1960:20), the "second specimen of Procariama" described by Rovereto (1914) is part of the same specimen that this author described as Phororhacos incertus, thus being added to the originally described type material (distal tarsometatarsus associated to the phalanges). The possibility of confusing material between these two species of similar sizes and the same geological age serves as a warning.
CONCLUSIONS
The family Phorusrhacidae belonging to the order Ralliformes, suborder Cariamae, consists of 13 genera and 17 species recognized nowadays, all being medium or large built birds, some of which even gigantic, rivalling with the largest already existing birds.
All are extinct. Their fossils are known as dating from the Middle Paleocene up to the limit Pliocene-Pleistocene (Fig. 34).
They certainly diversified at the end of the Cretaceous or beginning of the Cenozoic in South America, when this continental mass was insulated, separated from the remaining portions of the earth originating from the ancient supercontinent of Gondwana.They constituted an endemism very peculiar to South America, where they were the most spectacular land carnivores during most of the Tertiary. After South America became linked to North America, which occured with the raising of the isthmus of Panama, in the Upper Pliocene, the family also reached North America, where from at least one species is known (Titanis walleri).
Within the suborder Cariamae, the families Cariamidae, Idiornithidae and Bathornithidae are the closest to the Phorusrhacidae, and the identification of some fossil remains could be difficult if based on few segments of skeletons. Outstanding amongst the main characteristics of the family Phorusrhacidae are: the large or gigantic build; the narrow body, especially the upper maxilla, thorax and pelvis; the bulky and especially high upper maxilla, with the pointed, strong and curved apex; the existance of basipterigoid processes in the skull; the absence of uncinate processes on the ribs; the atrophiated cranial half of the pubis; the reduction of the wings and loss of the ability to fly; the extreme reduction of the acrocoracoid and procoracoid processes of the coracoid.
Those forms assigned to the family Phorusrhacidae from the Lower Tertiary of Europe and North America do not belong to this family; they present certain plesiomorphic similarities and are represented by only a few segments of the skeleton, which do not permit an adequate comparison, thus requiring a deeper revision in their phylogenetic afinities.
Moreover, some of the South American forms described based on only few segments of the skeleton, that do not allow for arriving at an effective conclusion as to their classification, even though several authors have already related them therein, are excluded from the family Phorusrhacidae. Included in this case are: Riacama caliginea Ameghino, 1899, Smiliornis penetrans Ameghino, 1899, Pseudolarus guaraniticus Ameghino, 1899, Pseudolarus eocaenus Ameghino, 1899 and Lophiornis obliquus Ameghino, 1891.
The proposed final classification is:
Order Ralliformes Reichenbach, 1852
Suborder Cariamae Fürbringer, 1888
Family Phorusrhacidae Ameghino, 1899
1 Subfamily Brontornithinae Moreno & Mercerat, 1891
Genus Brontornis Moreno & Mercerat, 1891
B. burmeisteri Moreno & Mercerat, 1891
Genus Physornis Ameghino, 1895
P. fortis Ameghino, 1895
Genus Paraphysornis Alvarenga, 1993
P. brasiliensis (Alvarenga, 1982)
2 Subfamily Phorusrhacinae Ameghino, 1889
Genus Phorusrhacos Ameghino, 1889
P. longissimus Ameghino, 1899
Genus Devincenzia Kraglievich, 1932
D. pozzi (Kraglievich, 1931)
Genus Titanis Brodkorb, 1963
T. walleri Brodkorb, 1963
3 Subfamily Patagornithinae Mercerat, 1897
Genus Patagornis Moreno & Mercerat, 1891
P. marshi Moreno & Mercerat, 1891
Genus Andrewsornis Patterson, 1941
A. abbotti Paterson, 1941
Genus Andalgalornis Patterson & Kraglievich, 1960
A. steulleti (Kraglievich, 1931)
4 Subfamily Psilopterinae Dolgopol de Saez, 1927
Genus Psilopterus Moreno & Mercerat, 1891
P. bachmanni (Moreno & Mercerat, 1891)
P. lemoinei (Moreno & Mercerat, 1891)
P. affinis (Ameghino, 1899)
P. colzecus Tonni & Tambussi, 1988
Genus Procariama Rovereto, 1914
P. simplex Rovereto, 1914
Genus Paleopsilopterus Alvarenga, 1985
P. itaboraiensis Alvarenga, 1985
5 Subfamily Mesembriornithinae Kraglievich, 1932
Genus Mesembriornis Moreno, 1889
M. milneedwardsi Moreno, 1889
M. incertus (Rovereto, 1914)
ACKNOWLEDGMENTS
This study was supported by grants from the National Geographic Society (3183-85 and 3699-87), and Coordenadoria de Apoio a Pesquisa e Ensino Superior (CAPES) (HMFA) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (EH). We are indebted to the authorities of the American Museum of Natural History; the Natural History Museum, London; the Divisão de Geologia e Mineralogia do Departamento Nacional da Produção Mineral, Rio de Janeiro; the Field Museum of Natural History; the Museo Argentino de Ciencias Naturales Bernardino Rivadavia; the Muséum National d'Histoire Naturelle, Paris; the Museo de La Plata; the Museo Nacional de História Natural de Montevideo; the Museu Nacional do Rio de Janeiro; and the Forschunginstitut Senckenberg, Frankfurt where this studies were accomplished. We thank Miguel Soria H., Luis M. Chiappe, Eduardo Tonni, Claudia Tambussi, Jorge Noriega, Rosendo Pascual, José F. Bonaparte and Julia Clark for some special discussions on phorusrhacids. We wish to thank Eduardo Parentoni Bretas for the reproduction of the life appearance of some phorusracids.
Recebido em 30.08.2002
Aceito em 09.10.2002
- Alvarenga, H. 1982. Uma gigantesca ave fóssil do Cenozóico brasileiro: Physornis brasiliensis sp. n. Anais da Academia Brasileira de Ciências, 54:697-712.
- Alvarenga, H. 1985a. Um novo Psilopteridae (Aves: Gruiformes) dos sedimentos Terciários de Itaboraí, Rio de Janeiro, Brasil. In: Congresso Brasileiro de Paleontologia, 8. Anais Rio de Janeiro, NME-DNPM, 1983. p.1720. (Série Geologia, 27)
- Alvarenga, H. 1985b. Notas sobre os Cathartidae (Aves) e descrição de um novo gênero do Cenozóico brasileiro. Anais da Academia Brasileira de Ciências, 57:349-357.
- Alvarenga, H. 1990. Flamingos fósseis da bacia de Taubaté, Estado de São Paulo, Brasil: descrição de nova espécie. Anais da Academia Brasileira de Ciências 62:335-345.
- Alvarenga, H. 1993. Paraphysornis novo gênero para Physornis brasiliensis Alvarenga, 1982 (Aves: Phorusrhacidae). Anais da Academia Brasileira de Ciências, 65:403-406.
- Amadon, D. 1947. An estimated weight of the largest known bird. Condor, 49:159-164.
- Ameghino, F. 1887. Enumeración sistemática de las espécies de mamíferos fósiles coleccionados por Carlos Ameghino en los terrenos Eocenos de la Patagonia austral y depositados en el Museo de La Plata. Boletim Museo La Plata, 1:1-26.
- Ameghino, F. 1889. Contribuición al conocimiento de los mamíferos fósiles de la República Argentina. Actas Academia Nacional Ciencias de Cordoba, 6:1-1028.
- Ameghino, F. 1891a. Mamíferos y aves fósiles Argentinos: espécies nuevas: adiciones y correciones. Revista Argentina Historia Natural, 1:240-259.
- Ameghino, F. 1891b. Enumeración de las aves fósiles de la República Argentina. Revista Argentina Historia Natural, 1:441-453.
- Ameghino, F. 1895. Sobre las aves fósiles de Patagonia. Boletin del Instituto Geografico da Argentina, 15:501-602.
- Ameghino, F. 1898. Sinopsis geológico-paleontológica de la Argentina. Segundo Censo de la República Argentina, 1:112-255.
- Ameghino, F. 1899. Sinopsis geológico-paleontológica. Suplemento (Adiciones y correciones), La Plata, 13p.
- Ameghino, F. 1901. L'age des formations sédimentaires de Patagonie. Anales de la Sociedad Cientifica Argentina, 51:65-91.
- Andrews, C. 1896. Remarks on the Stereornithes, a group of extinct birds from Patagonia. Ibis, 7:1-12.
- Andrews, C. 1899. On the extinct birds of Patagonia. Transactions of the Zoological Society of London, 15:55-86.
- Baskin, J.A. 1995. The giant flightless bird Titanis walleri (Aves: Phorusrhacidae) from the Pleistocene coastal plain of south Texas. Journal of Vertebrate Paleontology, 15:842-844.
- Baumel, J. &. Witmer, L. 1993. Osteologia. In: Baumel, J. ; King, A.; Breazile, J.; Evans, H. & Berge, J. (eds.), Nomina Anatomica Avium 2. ed., Cambridge. Nuttal Ornithological Club. p.45-132. (Publications of the Nuttall Ornithological Club 23)
- Brodkorb, P. 1963. A giant flightless bird from the Pleistocene of Florida. Auk, 80:111-115.
- Brodkorb, P. 1967. Catalogue of fossil birds, Part III (Ralliformes, Ichthyornithiformes, Charadriiformes). Bulletin of Florida State Museum, 2:99-220.
- Cabrera, A. 1939. Sobre vertebrados fósiles del Plioceno de Adolfo Alsina. Revista del Museo La Plata, 2:3-35.
- Campbell Jr., K. & Marcus, L. 1992. The relationship of hindlimb bone dimensions to body weight in birds. Science Series Natural History Museum of Los Angeles County, 36:395-412.
- Campbell Jr., K. &. Tonni, E. 1980. A new genus of teratorn from the Huayquerian of Argentina (Aves: Teratornithidae). Contributions in Science Natural History Museum of Los Angeles County, 330:59-68.
- Case, J.A.; Woodburne, M & Chaney, D. 1987. A gigantic phororhacoid (?) bird from Antarctica. Journal of Paleontology, 61:1280-1284.
- Carr, G.E. 1981. An early Pleistocene avifauna from Inglis Florida Gainesville, Univ. Florida, 161p. (Ph.D. Dissertation)
- Chandler, R.M. 1994. The wing of Titanis walleri (Aves: Phorusrhacidae) from the late Blancan of Florida. Bulletin of Florida Museum of Natural History, 36:175-180.
- Chiappe, L. & Soria, M. 1990. Phororhacos Ameghino, 1889 (Aves, Gruiformes): proposed conservation. Bulletin of Zoological Nomenclature, 3:198-201.
- Cracraft, J. 1968. A review of the Bathornithidae (Aves, Gruiformes), with remarks on the relationships of the suborder Cariamae. American Museum Novitates, 2326:1-46.
- Cracraft, J. 1971. Systematics and evolution of the Gruiformes (Class Aves) 2. Additional comments on the Bathornithidae, with descriptions of new species. American Museum Novitates, 2449:1-14.
- Cracraft, J. 1976. The species of moas (Aves: Dinornithidae). Smithsonian Contributions to Paleobiology, 27:189-205.
- Dolgopol de Saez, M. 1927. Las aves corredoras fósiles del Santacrucense. Anales de la Sociedad Cientifica Argentina, 103:145-64.
- Gilbert, B.; Martin, L. & Savage, H. 1981. Avian Osteology Laramie, B. Miles Gilbert Publisher. 252p.
- Howard, H. 1929. The avifauna of Emeryville shellmound. University of California Publications in Zoology, 32:301-394.
- Kraglievich, L. 1931. Contribución al conocimiento de las aves fósiles de la época araucoentrerriana. Physis, 10:304-315.
- Kraglievich, L. 1932. Una gigantesca ave fósil del Uruguay, Devincenzia gallinali n. gen. n. sp., tipo de una nueva familia, Devincenziidae, del Orden Stereornithes. Anales del Museo Historia Natural de Montevideo, 3:323-55.
- Kraglievich, L. 1946. Noticia preliminar acerca de un nuevo y gigantesco Estereornito de la fauna Chapadmalense. Anales de la Sociedad Cientifica Argentina, 142:104-121.
- Lambrecht, K. 1933. Handbuch der Paleornithologie Berlin, Gebrüder Bornthreger. 1024p.
- Livezey, B.C. 1998. A phylogenetic analysis of the Gruiformes (Aves) based on morphological characters, with an emphasis on the rails (Rallidae). Transactions of the Royal Society of London, 353:2077-2151.
- Loomis, F.B. 1914. The Deseado formation of Patagonia Massachussets, Public Amherst College. 232p.
- Lydekker, R. 1891. Catalogue of the fossil birds in the British Museum (Natural History) London, British Museum Publishers. 368p.
- Lydekker, R. 1893. On the extinct giant birds of Argentina. Ibis, 5:40-47.
- MacFadden, B.J. 1985. Essay review of Marshall, Hoffstetter and Pascual, drifting continents, mammals, and time scales: current developments in South America. Journal of Vertebrate Paleontology, 5:169-74.
- Marshall, L.G.; Cifelli, R.; Drake, R. & Curtis, G. 1986. Vertebrate paleontology, geology, and geochronology of the Tapera de Lopez and Scarritt Pocket, Chubut Province, Argentina. Journal of Paleontology, 60:920-951.
- Mercerat, A. 1893. Note sur la géologie de Patagonie Buenos Aires. 6p.
- Mercerat, A. 1897. Note sur les oiseaux fossiles de la Republique Argentine. Anales de la Sociedad Cientifica Argentina, 43:222-240.
- Moreno, F.P. 1889. Breve reseña de los progresos del Museo La Plata, durante el segundo semestre de 1888. Boletin del Museo La Plata, 3:1-44.
- Moreno, F.P. & Mercera, A. 1891. Catálogo de los pájaros fósiles de la República Argentina conservados en el Museo de La Plata. Anales del Museo de La Plata, 1:7-71.
- Mourer-Chauviré, C. 1981. Première indication de la présence de Phorusrhacidés, famille d'oiseaux géants d'Amérique du Sud, dans le Tertiaire Européen: Ameghinornis nov. gen. (Aves, Ralliformes) des Phosphorites du Quercy, France. Geóbios, 14:637-647.
- Mourer-Chauviré, C. 1983. Les Gruiformes (Aves) des Phosphorites du Quercy (France). I. Sous-ordre Cariamae (Cariamidae et Phorusrhacidae) systématique et biostratigraphie. Paleovertebrata, 13:83-143.
- Noriega, J. 1995. The avifauna from the "Mesopotamian" (Ituzaingó Formation; Upper Miocene) of Entre Rios Province, Argentina. Courier Forschungsinstitut Senckenberg, 181:141-148.
- Olson, S. 1985. The fossil record of birds. In: Farner, D.; King, J. & Parkes, K. (eds.). Avian Biology New York, Academic Press. v.8, p.79-252.
- Patterson, B. 1941. A new phororhacoid bird from the Deseado formation of Patagonia. Field Museum of Natural History, Geological Series, 8:49-54.
- Patterson, B. & Kraglievich, L. 1960. Sistemática y nomenclatura de las aves fororracoideas del Plioceno Argentino. Publicacion del Museo Municipal Ciencias Naturales y Tradicionales de Mar del Plata, 1:1-51.
- Peters, D.S. 1987. Ein "Phorusrhacidae" aus dem Mittel-Eozan von Messel (Aves: Gruiformes: Cariamae). Documents des Laboratoires de Géologie de Lyon, 99:71-87.
- Richmond, C. 1902. List of generic terms proposed for birds during the years 1890 to 1900, inclusive, to which are added names omitted by Waterhouse in his "Index generum avium". Proceedings United States National Museum, 24:663-730.
- Rovereto, C. 1914. Los estratos araucanos y sus fósiles. Anales del Museo Nacional Historia Natural de Buenos Aires, 25:1-247.
- Rusconi, C. 1946. Algunos mamíferos, reptiles y aves del Oligoceno de Mendoza. Revista de la Sociedad de Historia y Geografia de Cuyo, 2:1-37.
- Sinclair, W. & Farr, M. 1932. Aves of the Santa Cruz beds. In: Scott, W. (ed.) Reports of the Princeton University expeditions to Patagonia (1896-1899) Princeton University. v.7, p.157-191.
- Soria, M. & Alvarenga, H. 1989. Nuevos restos de mamíferos de la cuenca de Taubaté, Estado de São Paulo, Brasil. Anais da Academia Brasileira de Ciências, 61(2):157-175.
- Thiollay, J. 1994. Family Accipitridae (hawks and eagles). In: Hoyo, J.; Elliott, A. & Sargatal, J. (eds.), Handbook of the birds of the world Barcelona, Lynx Edicions. v.2, p.52-205.
- Tambussi, C.; Ubilla, M. &. Perea, D. 1999. The youngest large carnassial bird (Phorusrhacidae, Phorusrhacinae) from South America (Pliocene-Early Pleistocene of Uruguay). Journal of Vertebrate Paleontology, 19:404-406.
- Tonni, E. 1980. The present state of knowledge of the cenozoic birds of Argentina. Contributions in Science Natural History Museum of Los Angeles County, 330:105-114.
- Tonni, E. & Tambussi, C. 1988. Un nuevo Psilopterinae (Aves: Ralliformes) del Mioceno tardio de la Provincia de Buenos Aires, Republica Argentina. Ameghiniana, 25:155-160.
- Wetmore, A. 1944. A new terrestrial vulture from the Upper Eocene deposits of Wyoming. Annals of the Carnegie Museum, 30:57-69.
- Wetmore, A. 1967. Re-creating Madagascar's giant extinct bird. National Geographic, 132:488-493.
- Wroe, S. 1998. Bills, bones and bias: did thunder birds eat meat? Riversleigh Notes, 40:2-4.





Publication Dates
-
Publication in this collection
31 Oct 2003 -
Date of issue
2003
History
-
Received
30 Aug 2002 -
Accepted
09 Oct 2002