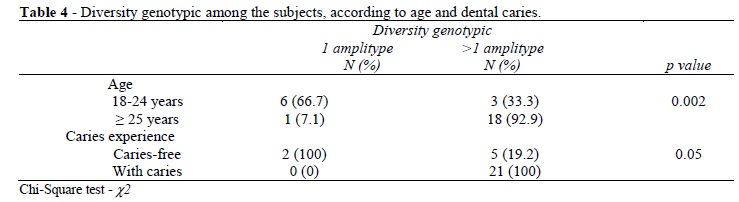Abstract
The objective of this study was to analyze the genotypic diversity, frequency of serotypes and the detection of mutacins from Streptococcus mutans isolates in caries-free and caries-active individuals.A total of 260 S. mutans isolated from 28 individuals with and without dental caries were subjected to AP-PCR and PCR screening of glucosyltransferase B, mutacin and serotype genes, which showed the presence of. 70 different genotypes. There was no statistically significant association between the presence of genes for serotypes and mutacins with dental caries. However, there was a statistically significant and a strong association between the higher genotypic diversity in the subjects with caries (r = 0.72, p = 0.001). There was an increase in the number of genotypes with increasing age (p <0.01).
Dental caries; genotypic diversity; mutacin; serotype; Streptococcus mutans
HUMAN AND ANIMAL HEALTH
Genotypic diversity and virulence factors of Streptococcus mutans in caries-free and caries-active individuals
Miula Portelinha BragaI; Augusta PiovesanI; Natália ValariniI; Sandra Mara MacielII; Flaviana Bombarda de AndradeIII; Regina Célia Poli-FredericoI,* * Author for correspondence: reginafrederico@yahoo.com.br
IUniversidade Norte do Paraná; Londrina - PR - Brasil
IIUniversidade Estadual de Maringá e Universidade Norte do Paraná; Londrina - PR - Brasil
IIIFaculdade de Odontologia de Bauru; Universidade de São Paulo; São Paulo - SP - Brasil
ABSTRACT
The objective of this study was to analyze the genotypic diversity, frequency of serotypes and the detection of mutacins from Streptococcus mutans isolates in caries-free and caries-active individuals.A total of 260 S. mutans isolated from 28 individuals with and without dental caries were subjected to AP-PCR and PCR screening of glucosyltransferase B, mutacin and serotype genes, which showed the presence of. 70 different genotypes. There was no statistically significant association between the presence of genes for serotypes and mutacins with dental caries. However, there was a statistically significant and a strong association between the higher genotypic diversity in the subjects with caries (r = 0.72, p = 0.001). There was an increase in the number of genotypes with increasing age (p <0.01).
Key words: Dental caries, genotypic diversity, mutacin, serotype, Streptococcus mutans
INTRODUCTION
Mutans streptococci are generally accepted as one of the principal aetiologic agents of dental caries. The dental biofilm consistes of a complex bacterial community and the ability of specific strains of Streptococcus mutans (S. mutans) to compete with other strains may be essential for colonization (Loesche 1986; Becker et al. 2002). S. mutans show high phenotypic and genotypic heterogeneities (Mattos-Graner et al. 2000, Mattos-Graner et al. 2001; Saarela et al. 1996) and some genotypes may be able to colonize the host and induce dental caries better than other genotypes. This hypothesis has been reinforced by the findings of significant variations in caries induction potential in the animal models when different S. mutans strains were compared (Köhler and Krasse 1990). In addition, variations in some virulence traits in the S. mutans isolates were associated to caries activity of the infected hosts and to their ability to grow as biofilms (Mattos-Graner et al. 2000; Mattos-Graner et al. 2001).
Most clones of S. mutans produce bacteriocins, named mutacins, which are active against oral streptococci, including other clones of S. mutans (Alaluusua et al. 1991), and also against non-related organisms, e.g. Neisseria spp. (Mota-Meira et al. 2000). The broad activity spectrum of some mutacins against distinct genotype of S. mutans highlights the important biotechnological potential of these substances in dental caries control (Hillman et al. 1987).
The classification of mutacin-producer strains based on their bactericidal activity, sensitivity to other self-produced mutacins and presence of plasmids, divides mutacins in four types, I, II, III and IV (Caufield et al. 1985; Morency et al. 1995; Qi et al. 1999a; Qi et al. 2001). The structural genes of the prepropeptides of mutacins I, II, III and IV (mutA) have been sequenced (Qi et al. 1999b; Qi et al. 2001), and their biosynthetic locus is formed by several genes, including those involved in regulation, cleavage, transport and immunity to the produced mutacin (Chen et al. 1999; Qi et al. 1999a).
S. mutans strains have been classified into c, e and f serotypes based on the chemical composition of their cell surface polysaccharides (Linzer et al. 1986). The serotype-specific polysaccharide of S. mutans is known to consist of rhamnose-glucose polymers, with a backbone of rhamnose and side chains of α- or ß-linked glucosidic residues. A serotype-specific polysaccharide of S. mutans was demonstrated to play important roles in streptococcal adherence to human monocytic and fibroblastic cells, and was speculated to be its most efficient cytokine-stimulating component (Engels-Deutsch et al. 2003). The serotype cS. mutans strains are predominant in the human oral cavity among the serotype c, e and f strains (Gronroos et al. 1995). The serotype c RGP structure may have advantages for S. mutans colonization of the oral cavity (Shibata et al. 2003).
The objective of the present study was to analyze the genetic diversity, frequency of serotype c, e and f, detection of mutacins I, II, III and IV from S. mutans isolates in caries-free and caries-active individuals.
MATERIALS AND METHODS
Subjects
The study sample consisted of 28 preschools aged 18 to 34 months, randomly selected, from a public Early Childhood Center in Londrina, southern Brazil. The aim and details of the experiments were explained and the informed consent forms were obtained prior to beginning the research procedure. The experimental procedures were approved by the institutional Ethical Committee of the University of North of Parana School of Dentistry and by the local Health and Education Authorities (PP/034/06).
Caries experience was measured by the DMFT (decayed, missing and filled teeth) index, according to the World Health Organization criteria (WHO, 1997). The clinical examination was performed by the same examiner (FJSP). The intra-examiner agreement was high (κ=0.92). The caries experience was dichotomized into two groups: caries-free (dmft= 0) and dental caries children (dmft>0). The dental examination was performed in a natural lighted with direct visual inspection and tactile sensation with a dental explorer and mouth mirror.
Bacterial strains and DNA extraction
S. mutans clinical isolates were obtained from the Mitis-Salivarius Agar with bacitracin and potassium telurite. Ten colonies resembling S. mutans from each patient were transferred to the brain heart infusion broth BHI (Difco, Detroit, USA) and incubated at 37ºC for 48h in an anaerobic jar. The DNA from 280 isolates were extracted by using a simple DNA preparation in which the cells were washed and boiled for 10 minutes with TE buffer (10mM Tris/HCL, 1 mM EDTA, pH8.0). The debris was pelleted and the supernatants were stored in a freezer at -80ºC until use.
PCR analyses
To confirm the identity of the strains, PCR reactions were carried out with the primers specific for gtfB, encoding glucosyltransferase B: Forward: 5'ACT ACA CTT TCG GGT GGC TTG G3' and Reverse: 5'CAG TAT AAG CGC CAG TTT CAT C3' (Invitrogen, São Paulo, Brazil) described by Oho et al. (2000), yielding an amplicon of 517bp. A total of 260 isolates were identified as S. mutans: 240 strains were included from 26 caries-active individuals and 20 strains from 2 caries-free individuals.
Each reaction consisted of 1.0µl template DNA (100ng/µL), 1.0 µM of each primer, 200 µM of each dNTP, 1X PCR buffer (200 mmol l-1 Tris-HCl pH 8.4; 500 mmol l-1 KCl), 1.5 mM MgCl2 and 1 U Taq DNA polymerase (Invitrogen, São Paulo, Brazil) in a total volume of 25 µl. The amplification reaction was performed in 30 cycles as follows: denaturation 95ºC for 30s, annealing at 59ºC for 30s, and extension at 72ºC for 1 min. One reference strain (ATCC 25175) was used as a positive control of S. mutans and distilled water was used as a negative control. Amplification products were analysed electrophoretically in 1% agarose gels using TBE buffer (89 mmol l-1 Tris borate, 89 mmol l-1 boric acid, 2 mmol l -1 EDTA; pH 8.0), stained with Sybr Safe (Invitrogen) and observed under UV light. A 100 bp DNA ladder (Invitrogen, São Paulo, Brazil) served as molecular-size marker in each gel. All reactions were repeated at least twice.
AP-PCR typing
The strains identified as S. mutans were genotyped. The genetic diversity of S. mutans isolates was analyzed by AP-PCR reactions. The sequences of the primers OPA 02 (5'TGCCGAGCTG 3') and OPA 13 (5'CAGCACCCAC3') were used. The PCR reactions were performed as follows: 1X PCR buffer (200 mmol l-1 Tris-HCl pH 8.4; 500 mmol l-1 KCl) with 3.5mM of MgCl2, 0.2mM of each dNTPs, 0.4mM of primers, 1U of Taq DNA polymerase and 1.0µL of DNA sample. The PCR conditions included 35 cycles of denaturation at 94oC for 1 min, annealing at 36oºC for 2 min, extension at 72oºC for 2min, with initial denaturation at 94oC for 5 min and a final extension at 72oC for 5 min. The eletrophoresis was carried out as described above; however, the amplification products were analysed in 2% agarose gel.
The individual AP-PCR amplicons were marked and the individual bands were analyzed by using the Dice coefficient (>95%). A dendrogram was constructed using the UPGMA cluster analysis with the aid of NTSYS (Numerical Taxonomy and Multivariate Analysis System) program (Exeter Software, Setauket, NY).
PCR screening of mutacin and serotype genes
The detection of genes encoding mutacin types I, II, III and IV was performed by the PCR using specific primers to each type (Table 1). The PCR mixture for mutacins consisted of 1X PCR buffer 10x, 2.5 mM MgCl2, 200 ρmol of each deoxynucleotide, 0.3 µM of each oligonucleotide primer, 1U of Taq DNA polymerase (Invitrogen) and 50ng of template DNA. After denaturation at 94 ºC for 5 min, a total of 30 PCR cycles were performed; each cycle consisted of 30s of denaturation at 92 ºC, 30s of annealing at 55 ºC, and 1 min of extension at 72 ºC; and the final extension 5 min at 72 ºC.
For the detection of serotypes c, e and f, a multiplex PCR was realized. Three sets of specifics primers (SC-F plus SC-R, SE-F plus SE-R, and SF-F plus SF-R) were used to identify S.mutans serotypes (Table 2). The PCR mixture (10 µl) consisted of 0.2 mM each deoxyribonucleotide triphosphate, 1X PCR buffer, 2mM MgCl2, 1U Taq DNA polymerase (Invitrogen), 0.5µlM concentration of each primer, and 50ng of template DNA. After denaturation at 96 ºC for 2 min, a total of 25 PCR cycles were performed; each cycle consisted of 15s of denaturation at 96 ºC, 30s of annealing at 61 ºC, and 1 min of extension at 72 ºC.
The PCR products were analyzed by electrophoresis in 1% agarose gel using Tris/borate/EDTA buffer (pH 8-0). A 250 bp DNA ladder was included in each gel (Invitrogen). The DNA was stained with 0.5 µg ml-1 ethidium bromide and visualized under UV illumination.
Statistical analysis
The chi-square test and the Spearman's coefficient of correlation were applied to analyze the associations and correlations between the genotypic diversity, differences in the frequency of mutacin and serotype genes and experience of dental caries. Statistical significance was considered to be at α<0.05. The software Statistical Package for Social Science, v. 17 (SSPS, Chicago, IL, USA) was used for the data analysis.
RESULTS
The clinical S. mutans were isolated from the individuals aged 18 34 years (mean ± SD, 28.6 ± 6.6). The prevalence of dental caries (DMFT > 0) was 92.9%, with a mean DMFT score of 11.64 (SD = 6.33). A total of 70 different genotypes were detected among the 260 S. mutans isolates. The test was performed with the OPA-02 and OPA-13, and each of these primers generated a different spectrum of amplicons, indicative of genetic polymorphism; when the results obtained with the two primers were combined, the strains were classified into two distinct genotypes in the caries-free and into 68 distinct genotypes from the caries subjects (Table 3). The comparison of the similarity indices showed greater diversity among the isolates from different individuals.
Considering the whole population, some of the subjects harbored just one genotype whereas others exhibited up to five genotypes. Seven subjects harbored one genotypic only and twenty-one had more than one genotype. Of these seven, two were caries-free and five had dental caries (Table 3). The analysis between the genotypic diversity and the caries experience showed a higher genotypic diversity in the subjects with caries (χ2= 6.46; p = 0.05) (Table 4). The Spearman correlation test showed a strong association between the genotypic diversity and caries experience (r = 0.72; P < .001).
Table 4 showed an increase in the number of genotypes with increasing age. The 66.7% of individuals aged 18 to 24 years had only one genotype, while 92.9% of individuals older than 25 years showed a higher genotypic diversity of S. mutans in their oral cavity. This association was statistically significant (χ2=12,38; p= 0.002). All the participants, except three, subject harbored serotype c isolates and serotype f was the next most common and the presence of serotype e was not identified (Table 3). The PCR screening with the primers of mutacin I, II and IV revealed that the isolates were positive for these mutacins in the caries-free subjects. The PCR for the mutacin III did not yield amplicon in any S. mutans isolates in this group (Table 3). The PCR with the primers of mutacin IV showed that 9 out of 28 (32.1%) S. mutans isolates were positive in the caries-active group; on the other hand, the amplicons I/III genes revealed that 6 out of 28 (21.4%) isolates carried these genes (Table 3).This study showed no statistically significant association between the presence of the genes for serotypes and mutacins with dental caries.
DISCUSSION
In the oral ecosystem, the development of the bacterial community generally involves a sucession of populations and competition for the receivers of adhesion, foods, and the production of inhibitory substances such as the bacteriocins (Marcotte and Lavoie 1998). The mutacins and serotypes have been implicated as the virulence factors in dental caries. The relationship between the caries activity and the higher synthesis of some virulence factors by different genotypes of S. mutans has been demonstrated in the literature (Napimoga et al. 2005). In the present study, the genotypic diversity of S. mutans was statistically associated to the dental caries. Preschools with dental caries showed more diversity than those without dental caries. This finding was contrary to the results obtained by Kreulen et al. (1997), who showed a negative relationship between the caries activity and genotype diversity. However, the present result was in agreement with earlier reports (Hirose et al. 1993; Alaluusua et al. 1996; Emanuelsson et al. 2003; Napimoga et al. 2004). It has been observed that the subjects harbor one to five distinct genotypes of S. mutans at different ages (Alallusuua et al. 1996; Emmanuelsson et al. 1998; Grönroos et al. 1998; Klein et al. 2004; Mattos-Graner et al. 2001; Caulfield and Walker 1989; Kulkarni et al. 1989; Kreulen 1997; Napimoga et al. 2004). In the population examined, two individuals had only one genotype in the group caries-free, and only two had as many as five genotypes in the dental caries group.
In a study of young adults, Emanuelsson et al. (2003) found a maximum of seven genotypes in the subjects who had previous experience of caries. Napimoga et al. (2004) also found a maximum of eight genotypes in the caries-active subjects using AP-PCR. Heavy colonization and growth of multiple genotypes in the same oral cavity are likely to be consequences of frequent consumption of fermentable carbohydrates. Different clonal types of S. mutans detected within the oral cavity of one subject can have different phenotype and genetic properties (Guo et al. 2006). The high clonal diversity of S. mutans is likely to result in colonization by the clones of different virulence (Caulfield 1997).
In relation to the frequency of mutacin, no association with dental caries could be found in this study. However, a higher proportion for mutacin I/III and to mutacin IV in S. mutans isolates from caries-active individuals was observed. Kamiya et al (2005) found that the isolates recovered from the caries-active individuals showed a higher frequency of detection of mutacins I/III and IV. Clinically, mutacins have been considered important for the establishment and equilibrium of bacteria in dental biofilms (Hillman et al. 1987). Supporting this hypothesis, the antimicrobial spectrum of mutacin IV is specifically against the members of the mitis group of oral streptococci (Qi et al. 2001). Nevertheless, the results showed that given the increasing complexity of the oral microbiota, as found in the caries-active individuals (Napimoga et al. 2004), the S. mutans strains producing a wide spectrum of mutacins, including mutacins I, II and III, could become prevalent in most oral sites.
The caries-active individuals, the sites from which S.mutans were isolated, showed higher diversity, probably because the production of organic acids and mutacins with the biofilm resulted in a more complex community compared to caries-free individuals (Paddick et al. 2003). Probably due to this complexity, S.mutans genotypes isolated from the caries-active individuals presented higher frequencies of mutacin IV and a wide spectrum of mutacins, such as I/III, and presented greater mutacin activity in vitro compared to S. mutans recovered from the caries-free individuals (Kamiya et al. 2005).
Qi et al. (2001) studied the clinical isolates of S. mutans for the presence of mutacin IV genes by the PCR and found > 50% positive results. The mutacin IV was produced by planktonic cells while mutacin I was produced by the biofilm-like cells. Different mutacins could serve different purposes during the process of colonization by S. mutans. For instance, the production of mutacin IV by the planktonic cells in saliva might help S. mutans kill the primary colonizers on the tooth surface to make the room for its own population. Supporting this hypothesis, the antimicrobial spectrum of mutacin IV has been shown to be specifically against members of the mitis group of oral streptococci.
Shibata et al. (2003) isolated S. mutans from 198 of 432 preschoool children (3 to 4 years old). The data revealed that serotype c predominated, serotype e was the next most commom, and serotype f occurred rarely in Japanese preschool children. This result was similar to the present results and with Gronroos et al. (1995). Furthermore, which found that the serotype f was the next most common and the presence of serotype e was not identified. It was noteworthy that no statistical association between the serotypes and dental caries was detected.
The findings of the present study point with respect to the higher genotypic diversity in the subjects with dental caries and higher frequency of serotype c and the genes for mutacins I, III and IV in the studied population.
ACKNOWLEDGMENTS
This study was supported by the grants from the FUNADESP.
REFERENCES
Alaluusua, S, Mttö , Grönroos L, Innila S, Torkko H, Asikainen S, et al. Oral colonization by more than one clonal type of mutans streptococcus in children with nursing-bottle dental caries. Arch Oral Biol. 1996; 41:167-173.
Becker MR, Paster BJ, Leys EJ, Moeschberger ML, Kenyon SG, Galvin Jamie L, et al. Molecular Analysis of Bacterial Species Associated with Childhood Caries. J Clin Microbiol. 2002; 40: 1001-1009.
Caufield PW, Childers NK, Allen DN, Hansen JB. Distinct bacteriocin groups correlate with different groups of Streptococcus mutans plasmids. Infect Immun. 1985; 48: 5156.
Caufield, PW. Dental caries a transmissible and infectious disease revisited: a position paper. Pediatr Dent. 1997; 19: 491-497.
Caufield PW, Walker TM. Genetic diversity within Streptococcus mutans evident fromchromosomal DNA restriction fragment polymorphisms. J Clin Microbiol. 1989; 27: 274278.
Chen P, Qi F, Novak J, Caufield PW. The specific genes for lantibiotic mutacin II biosynthesis in Streptococcus mutans T8 are clustered and can be transferred en bloc. Appl Environ Microbiol. 1999; 65: 1356-1360.
Emanuelsson I.M, Carlsson P, Hamberg K, Bratthall D. Tracing genotypes of mutans streptococci on tooth sites by random amplified polymorphic DNA (RAPD) analysis. Oral Microbiol Immunol. 2003; 18: 2429.
Engels-Deutsch M, Pini A, Yamashita Y, Shibata Y, Haikel Y, Scholler-Guinard M, et al. Insertional inactivation of pac and rmlB genes reduces the release of tumor necrosis factor alpha, interleukin-6, and interleukin-8 induced by Streptococcus mutans in monocytic, dental pulp, and periodontal ligament cells. Infect Immun. 2003; 71: 51695177.
Guo LH, Shi JN, Zhang Y, Liu XD, Duan J, Wei S. Identification of genetic differences between two clinical isolates of Streptococcus mutans by suppression subtractive hyridization. Oral Microbiol Immunol. 2006; 21: 372380.
Grönroos L, Mättö J, Saarela M, Luoma AR, Luoma H, Jousimies-Somer H, et al. Chorhexidine susceptibilities of mutans streptococcal serotypes and ribotypes. Antimicrob Agents Chemother. 1995; 39: 894-898.
Hillman JD, Dzuback AL, Andrews SW. Colonization of the human oral cavity by a Streptococcus mutans mutant producing increased bacteriocin. J Dent Res. 1987; 66: 1092-1094.
Hirose H, Hirose K, Isogai E, Miura H, Ueda I. Close association between Streptococcus sobrinus in the saliva of young children and smooth-surface caries increment. Caries Res. 1993; 27: 292297.
Kamiya RU, Napimoga MH, Rosa RT, Höfling JF, Gonçalves RB. Mutacins productin in Streptococcus Mutans genotypes isolated from caries-active and caries-free individuals. Oral Microbiol Immunol, 2005; 20: 20-24.
Klein MI, Florio FM, Pereira AC, Höfling JF, Gonçalves RB. Longitudinal study of transmission, diversity and stability of Streptococcus mutans and Streptococcus sobrinus genotypes in Brazilian nursery children. J Clin Microbiol. 2004; 42: 46204626.
Köhler B, Krasse B. Human strains of mutans streptococci show different cariogenic potential in the hamster model. Oral Microbiol Immunol. 1990; 5: 177-180.
Kreulen CM, de Soet HJ, Hogeveen R, Veerkamp JS. Streptococcus mutans in children using nursing bottles. J Dent Children. 1997; 64: 107111.
Kulkarni GV, Chan KH, Sandham HJ. An investigation into the use of restriction endonuclease analysis for the study of transmission of mutans streptococci. J Dental Res. 1989; 68: 11551161.
Linzer R, Reddy MS, Levine MJ. Immunochemical aspects of serotype carbohydrate antigen of Streptococcus mutans. In: Hamada S, Michalek S, Kiyono H, Manaker L, McGhee JR. Molecular Microbiology and Immunology of Streptococcus mutans. Amsterdam: Elsevier Science Publishers; 1986. pp. 2938.
Loesche WJ. Role of Streptococcus mutans in human dental decay. Microbiol Rev. 1986; 50: 353380.
Marcotte H, Lavoie MC. Oral microbial ecology and the role of salivary immunoglobulin A. Microbiol Mol Biol Rev. 1998; 62: 71-109.
Mattos-Graner RO, Smith DJ, King WF, Mayer MP. Water-insoluble glucan synthesis by mutans streptococcal strains correlates with caries incidence in 12- to 30-month-old children. J Dent Res., 2000; 79: 1371-1377.
Mattos-Graner RO, Li Y, Caufield PW, Duncan M, Smith DJ. Genotypic diversity of mutans streptococci in Brazilian nursery children suggests horizontal transmission. J Clin Microbiol. 2001; 39: 2313-2316.
Mota-Meira M, LaPointe G, Lacroix C, Lavoie MC. MICs of mutacin B-Ny266, nisin A, vancomycin, and oxacillin against bacterial pathogens. Antimicrob Agents Chemother. 2000; 44: 2429.
Morency H, Trahan L, Lavoie MC. Preliminary grouping of mutacins. Can J Microbiol, 1995; 41: 826831.
Napimoga MH, Kamiya RU, Rosa RT, Rosa EA, Höfling JF, Mattos-Graner RO, et al. Genotypic diversity and virulence traits of Streptococcus mutans in caries-free and caries-active individuals. J Med Microbiol. 2004; 53: 697-703.
Novak J, Caufield PW, Miller EJ. Isolation and biochemical characterization of a novel lantibiotic mutacin from S. Mutans. J Bacteriol. 1994; 176: 316 4320.
Oho T, Yamashita Y, Shimazaki Y, Kushiyama M, Koga T. Simple and rapid detection of Streptococcus mutans and Streptococcus sobrinus in human saliva by polymerase chain reaction. Oral Microbiol Immunol, 2000; 15: 258-262.
Paddick JS, Brailsford SR, Kidd EAM, Gilbert SC, Clark DT, Alam S, et al. Effect of the environment on genotypic diversity of Actinomyces naeslundii and Streptococcus oralis in the oral biofilm. Appl Environ Microbiol. 2003; 69: 6475-6480.
Qi F, Chen P, Caufield PW. Functional analyses of the promoters in the lantibiotic mutacin II biosynthetic locus in Streptococcus mutans. Appl Environ Microbiol, 1999a; 65: 652-658.
Qi F, Chen P, Caufield PW. Purification of the mutacin III from group III Streptococcus mutans UA 787 and genetic analyses of mutacin III biosynthesis genes. Appl Environ Microbiol. 1999b; 65: 3880-3887.
Qi F, Chen P, Caufield PW. Purification and biochemical characterization of mutacin I from the group I strain of Streptococcus mutans, CH43, and genetic analysis of mutacin I biosynthesis genes. Appl Environ Microbiol. 2000; 66: 3221-3229.
Qi F, Chen P, Caufield PW. The group I strain of Streptococcus mutans, UA140, produces both the lantibiotic mutacin I and a nonlantibiotic bacteriocin, mutacin IV. Appl Environ Microbiol. 2001; 67: 15-21.
Saarela M, Hannula J, Matto J, Asikainen S, Alaluusua S. Typing of mutans streptococci by arbitrarily primed polymerase chain reaction. Arch. Oral Bio. 1996; 41: 821826.
Shibata Y, Ozaki K, Seki M, Kawato T, Tanaka H, Nakano Y. Analysis of loci required for determination of serotype antigenicity in Streptococcus mutans and its clinical utilization. I Clin Microbiol. 2003; 41: 4107-4112.
World Health Organization, Oral Health Surveys Basic Methods, WHO, Geneva, Switzerland, 4th edition, 1997.
Received: November 29, 2011;
Revised: March 22, 2012;
Accepted: September 21, 2012.
- Alaluusua, S, Mttö , Grönroos L, Innila S, Torkko H, Asikainen S, et al. Oral colonization by more than one clonal type of mutans streptococcus in children with nursing-bottle dental caries. Arch Oral Biol. 1996; 41:167-173.
- Becker MR, Paster BJ, Leys EJ, Moeschberger ML, Kenyon SG, Galvin Jamie L, et al. Molecular Analysis of Bacterial Species Associated with Childhood Caries. J Clin Microbiol. 2002; 40: 1001-1009.
- Caufield PW, Childers NK, Allen DN, Hansen JB. Distinct bacteriocin groups correlate with different groups of Streptococcus mutans plasmids. Infect Immun. 1985; 48: 5156.
- Caufield, PW. Dental caries a transmissible and infectious disease revisited: a position paper. Pediatr Dent 1997; 19: 491-497.
- Caufield PW, Walker TM. Genetic diversity within Streptococcus mutans evident fromchromosomal DNA restriction fragment polymorphisms. J Clin Microbiol. 1989; 27: 274278.
- Chen P, Qi F, Novak J, Caufield PW. The specific genes for lantibiotic mutacin II biosynthesis in Streptococcus mutans T8 are clustered and can be transferred en bloc. Appl Environ Microbiol 1999; 65: 1356-1360.
- Emanuelsson I.M, Carlsson P, Hamberg K, Bratthall D. Tracing genotypes of mutans streptococci on tooth sites by random amplified polymorphic DNA (RAPD) analysis. Oral Microbiol Immunol. 2003; 18: 2429.
- Engels-Deutsch M, Pini A, Yamashita Y, Shibata Y, Haikel Y, Scholler-Guinard M, et al. Insertional inactivation of pac and rmlB genes reduces the release of tumor necrosis factor alpha, interleukin-6, and interleukin-8 induced by Streptococcus mutans in monocytic, dental pulp, and periodontal ligament cells. Infect Immun. 2003; 71: 51695177.
- Guo LH, Shi JN, Zhang Y, Liu XD, Duan J, Wei S. Identification of genetic differences between two clinical isolates of Streptococcus mutans by suppression subtractive hyridization. Oral Microbiol Immunol. 2006; 21: 372380.
- Grönroos L, Mättö J, Saarela M, Luoma AR, Luoma H, Jousimies-Somer H, et al. Chorhexidine susceptibilities of mutans streptococcal serotypes and ribotypes. Antimicrob Agents Chemother 1995; 39: 894-898.
- Hillman JD, Dzuback AL, Andrews SW. Colonization of the human oral cavity by a Streptococcus mutans mutant producing increased bacteriocin. J Dent Res. 1987; 66: 1092-1094.
- Hirose H, Hirose K, Isogai E, Miura H, Ueda I. Close association between Streptococcus sobrinus in the saliva of young children and smooth-surface caries increment. Caries Res. 1993; 27: 292297.
- Kamiya RU, Napimoga MH, Rosa RT, Höfling JF, Gonçalves RB. Mutacins productin in Streptococcus Mutans genotypes isolated from caries-active and caries-free individuals. Oral Microbiol Immunol, 2005; 20: 20-24.
- Klein MI, Florio FM, Pereira AC, Höfling JF, Gonçalves RB. Longitudinal study of transmission, diversity and stability of Streptococcus mutans and Streptococcus sobrinus genotypes in Brazilian nursery children. J Clin Microbiol. 2004; 42: 46204626.
- Köhler B, Krasse B. Human strains of mutans streptococci show different cariogenic potential in the hamster model. Oral Microbiol Immunol 1990; 5: 177-180.
- Kreulen CM, de Soet HJ, Hogeveen R, Veerkamp JS. Streptococcus mutans in children using nursing bottles. J Dent Children. 1997; 64: 107111.
- Kulkarni GV, Chan KH, Sandham HJ. An investigation into the use of restriction endonuclease analysis for the study of transmission of mutans streptococci. J Dental Res. 1989; 68: 11551161.
- Linzer R, Reddy MS, Levine MJ. Immunochemical aspects of serotype carbohydrate antigen of Streptococcus mutans In: Hamada S, Michalek S, Kiyono H, Manaker L, McGhee JR. Molecular Microbiology and Immunology of Streptococcus mutans. Amsterdam: Elsevier Science Publishers; 1986. pp. 2938.
- Loesche WJ. Role of Streptococcus mutans in human dental decay. Microbiol Rev. 1986; 50: 353380.
- Marcotte H, Lavoie MC. Oral microbial ecology and the role of salivary immunoglobulin A. Microbiol Mol Biol Rev. 1998; 62: 71-109.
- Mattos-Graner RO, Smith DJ, King WF, Mayer MP. Water-insoluble glucan synthesis by mutans streptococcal strains correlates with caries incidence in 12- to 30-month-old children. J Dent Res, 2000; 79: 1371-1377.
- Mattos-Graner RO, Li Y, Caufield PW, Duncan M, Smith DJ. Genotypic diversity of mutans streptococci in Brazilian nursery children suggests horizontal transmission. J Clin Microbiol. 2001; 39: 2313-2316.
- Mota-Meira M, LaPointe G, Lacroix C, Lavoie MC. MICs of mutacin B-Ny266, nisin A, vancomycin, and oxacillin against bacterial pathogens. Antimicrob Agents Chemother. 2000; 44: 2429.
- Morency H, Trahan L, Lavoie MC. Preliminary grouping of mutacins. Can J Microbiol, 1995; 41: 826831.
- Napimoga MH, Kamiya RU, Rosa RT, Rosa EA, Höfling JF, Mattos-Graner RO, et al. Genotypic diversity and virulence traits of Streptococcus mutans in caries-free and caries-active individuals. J Med Microbiol. 2004; 53: 697-703.
- Novak J, Caufield PW, Miller EJ. Isolation and biochemical characterization of a novel lantibiotic mutacin from S. Mutans. J Bacteriol 1994; 176: 316 4320.
- Oho T, Yamashita Y, Shimazaki Y, Kushiyama M, Koga T. Simple and rapid detection of Streptococcus mutans and Streptococcus sobrinus in human saliva by polymerase chain reaction. Oral Microbiol Immunol, 2000; 15: 258-262.
- Paddick JS, Brailsford SR, Kidd EAM, Gilbert SC, Clark DT, Alam S, et al. Effect of the environment on genotypic diversity of Actinomyces naeslundii and Streptococcus oralis in the oral biofilm. Appl Environ Microbiol 2003; 69: 6475-6480.
- Qi F, Chen P, Caufield PW. Functional analyses of the promoters in the lantibiotic mutacin II biosynthetic locus in Streptococcus mutans. Appl Environ Microbiol, 1999a; 65: 652-658.
- Qi F, Chen P, Caufield PW. Purification of the mutacin III from group III Streptococcus mutans UA 787 and genetic analyses of mutacin III biosynthesis genes. Appl Environ Microbiol 1999b; 65: 3880-3887.
- Qi F, Chen P, Caufield PW. Purification and biochemical characterization of mutacin I from the group I strain of Streptococcus mutans, CH43, and genetic analysis of mutacin I biosynthesis genes. Appl Environ Microbiol. 2000; 66: 3221-3229.
- Qi F, Chen P, Caufield PW. The group I strain of Streptococcus mutans, UA140, produces both the lantibiotic mutacin I and a nonlantibiotic bacteriocin, mutacin IV. Appl Environ Microbiol 2001; 67: 15-21.
- Saarela M, Hannula J, Matto J, Asikainen S, Alaluusua S. Typing of mutans streptococci by arbitrarily primed polymerase chain reaction. Arch. Oral Bio. 1996; 41: 821826.
- Shibata Y, Ozaki K, Seki M, Kawato T, Tanaka H, Nakano Y. Analysis of loci required for determination of serotype antigenicity in Streptococcus mutans and its clinical utilization. I Clin Microbiol 2003; 41: 4107-4112.
- World Health Organization, Oral Health Surveys Basic Methods, WHO, Geneva, Switzerland, 4th edition, 1997.
Publication Dates
-
Publication in this collection
24 May 2013 -
Date of issue
Apr 2013
History
-
Received
29 Nov 2011 -
Accepted
21 Sept 2012 -
Reviewed
22 Mar 2012