ABSTRACT
When confronted by predators, prey need to make an economic decision between continuing their current activity or flee. Flight Initiation Distance (FID), the distance at which an organism begins to flee an approaching threat, has been used to indicate an animal's fearfulness level and a way to examine factors influencing escape decisions. Here we investigated how the FID of the barber surgeonfish, Acanthurus bahianus, responds to the presence of spearfishers in a fishing site in northeast Brazil. Specifically, we examined whether the FID was influenced by body and group size; by the heterogeneity of species in groups formation; and the distance to shelter. Significant differences in FID were observed with increasing body size. We found no significant relationship of FID with size or group formation, neither with distance to shelter. Preferences in forming groups with A. bahianus were seen among some species, and a higher FID was associated with less sheltered substrates. Results obtained here support theories suggesting that spearfishers do influence fish behavior. We highlight that future research should focus on the indirect impacts of spearfishing on the structure of marine communities, emphasizing the anti-predator behaviour of juvenile and adult target fishes.
Keywords:
Antipredator behaviour; Brazil; Flight initiation distance; Rocky reefs; Spearfishing
RESUMO
Ao serem confrontadas por predadores, as presas precisam tomar decisões econômicas, escolhendo entre continuar com sua atividade atual ou fugir. Distância Inicial de Fuga (DIF), a distância à qual um organismo começa a fugir de uma ameaça que se aproxima, tem sido utilizada para indicar o nível de medo do animal e um modo para se examinar fatores que influenciam nas decisões de fuga. Aqui nós investigamos como a DIF do peixe cirurgião, Acanthurus bahianus , responde à presença de pescadores subaquáticos em uma tradicional área de pesca no nordeste do Brasil. Especificamente, investigamos se a DIF foi influenciada pelo tamanho do corpo e do grupo; pela heterogeneidade das espécies na formação de grupos; e pela distância para refúgio. Diferenças significativas na DIF foram observadas com o aumento no tamanho do corpo. Nós não encontramos nenhuma relação significativa da DIF com o tamanho ou formação do grupo, nem com a distância para refúgio. Preferências na formação de grupos com A. bahianus foram observadas entre algumas espécies,e maiores DIF foram associadas com substratos menos abrigados. Resultados obtidos aqui suportam teorias sugerindo que pescadores subaquáticos influenciam no comportamento dos peixes. Nós destacamos que pesquisas futuras devem focar nos impactos indiretos da pesca subaquática na estrutura das comunidades marinhas, enfatizando o comportamento anti-predador de peixes juvenis e adultos alvo da pesca.
Introduction
Risk perception in animals can be a result of the evolution of prey defensive traits in response to predator hunting behaviour (Abrams, 2000Abrams, P. A. 2000. The evolution of predator-prey interactions: theory and evidence. Annual Review of Ecology and Systematics, 31: 79-105.). Prey may adaptively respond to predators by becoming more vigilant and loosing foraging opportunities; remaining inactive and increasing the risk of capture (Brown et al., 2001Brown, J. S., B. P. Kotler & A. Bouskila. 2001. Ecology of fear: foraging games between predators and prey with pulsed resources. Annales Zoologici Fennici,38: 71-87.), or by moving away (flee) from an approaching threat (Dill, 1974Dill, L. M. 1974. The escape response of the zebra danio (Brachydanio rerio ) II. The effect of experience. Animal Behaviour, 22: 723-730.). Additionally, in making decisions toward flee or avoiding predators, prey can use their learning acquired with social transmitted information when responding to possible threats (Brown & Laland, 2003Brown, C. & K. N. Laland. 2003. Social learning in fishes: a review. Fish and Fisheries, 4: 280-288.; Kelley & Magurran, 2003Kelley, J. L. & A. E. Magurran. 2003. Learned predator recognition and antipredator responses in fishes. Fish and Fisheries, 4: 216-226.). Thereby, animals that are target by predators can adjust their antipredator behaviour responses, and consequently their flight decisions, according with their first perceptions of a probable risk (Ydenberg & Dill, 1986Ydenberg, R. C. & L. M. Dill. 1986. The Economics of fleeing from predators. Advances in the Study of Behavior, 16: 229-249.).
The main metric used to test risk perception and escape responses in wild animals is known as Flight Initiation Distance or FID. This index estimates the shorter distance at which the prey begins to flee from the predator approach (Ydenberg & Dill, 1986Ydenberg, R. C. & L. M. Dill. 1986. The Economics of fleeing from predators. Advances in the Study of Behavior, 16: 229-249.; Cooper & Frederick, 2007Cooper, W. E., Jr. & W. G. Frederick. 2007. Optimal flight initiation distance. Journal of Theoretical Biology, 244: 59-67.). FID has been used to examine the vigilance levels in a number of taxa, including birds (Blumstein, 2006Blumstein, D. T. 2006. Developing an evolutionary ecology of fear: how life history and natural history traits affect disturbance tolerance in birds. Animal Behaviour, 71: 389-399.), ungulates (Stankowich, 2008Stankowich, T. 2008. Ungulate flight responses to human disturbance: a review and meta-analysis. Biological Conservation, 141: 2159-2173.), lizards (Cooper, 2009Cooper, W. E., Jr. 2009. Flight initiation distance decreases during social activity in lizards (Sceloporus virgatus ). Behavioral Ecology and Sociobiology, 63: 1765-1771.), anurans (Bateman & Fleming, 2014Bateman, P. W. & P. A. Fleming. 2014. Living on the edge: effects of body size, group density and microhabitat selection on escape behaviour of southern leopard frogs Lithobates sphenocephalus . Current Zoology, 60: 712-718.), and fishes, which have been specially studied in the context of fishing (Feary et al., 2011Feary, D. A., J. E. Cinner, N. A. J. Graham & F. A. Januchowski-Hartley. 2011. Effects of customary marine closures on fish behavior, spear-fishing success, and underwater visual surveys. Conservation Biology, 25: 341-349.; Januchowski-Hartley et al., 2012Januchowski-Hartley, F. A., K. L. Nash & R. J. Lawton. 2012. Influence of spear guns, dive gear and observers on estimating fish flight initiation distance on coral reefs. Marine Ecology Progress Series, 469: 113-119.).
Variations in life history traits such as reproductive value, age at maturity and body size can affect the antipredator behaviour, and consequently how species will escape from particular threats (Blumstein, 2006Blumstein, D. T. 2006. Developing an evolutionary ecology of fear: how life history and natural history traits affect disturbance tolerance in birds. Animal Behaviour, 71: 389-399.). Large-bodied species might be more vulnerable, since they are generally less agile (Witter et al., 1994Witter, M. S., I. C. Cuthill & R. H. C. Bonser. 1994. Experimental investigations of mass-dependent predation risk in the European starling, Sturnus vulgaris . Animal Behaviour, 48: 201-222.), and so could have higher detection abilities, which may select for greater flight initiation distance (Blumstein et al ., 2005Blumstein, D. T., E. Fernández-Juricic, P. A. Zollner & S. C. Garity. 2005. Inter-specific variation in avian responses to human disturbance. Journal of Applied Ecology, 42: 943-953.; Blumstein, 2006Blumstein, D. T. 2006. Developing an evolutionary ecology of fear: how life history and natural history traits affect disturbance tolerance in birds. Animal Behaviour, 71: 389-399.). Likewise, predator characteristics (e.g. approach speed and predator size) are predicted to increase prey flight distances (Cooper, 2006Cooper, W. E., Jr. 2006. Dynamic risk assessment: prey rapidly adjust flight initiation distance to changes in predator approach speed. Ethology, 112: 858-864.; Stankowich & Coss, 2007Stankowich, T. & R. G. Coss. 2007. Effects of risk assessment, predator behavior, and habitat on escape behavior in Columbian black-tailed deer. Behavioral Ecology, 18: 358-367.; Cooper & Stankowich, 2010Cooper, W. E., Jr. & T. Stankowich. 2010. Prey or predator? Body size of an approaching animal affects decisions to attack or escape. Behavioral Ecology, 21: 1278-1284.). However, other variables may also modulate the escape performance to avoid predation in fishes, including physiology/biomechanics and behavioural ecology (Domenici et al., 2007Domenici, P., C. Lefrançois & A. Shingles. 2007. Hypoxia and the antipredator behaviours of fishes. Philosophical Transactions of the Royal Society B: Biological Sciences, 362: 2105-2121.; Domenici, 2010Domenici, P. 2010. Context-dependent variability in the components of fish escape response: integrating locomotor performance and behavior. Journal of Experimental Zoology Part A: Ecological Genetics and Physiology, 313: 59-79.; Marras et al., 2011Marras, S., S. S. Killen, G. Claireaux, P. Domenici & D. J. McKenzie. 2011. Behavioural and kinematic components of the fast-start escape response in fish: individual variation and temporal repeatability. Journal of Experimental Biology, 214: 3102-3110.).
Group size or distance to shelter, which are factors related to perceived risks and the cost of escaping, may also result in different effects on prey flight decisions (Stankowich & Blumstein, 2005Stankowich, T. & D. T. Blumstein. 2005. Fear in animals: a meta-analysis and review of risk assessment. Proceedings of the Royal Society B: Biological Sciences, 272: 2627-2634.). Fishes in shoals, taking advantage of the vigilance of other group members, might avoid predators to close approach, which can result in greater FID (Januchowski-Hartley et al., 2011Januchowski-Hartley, F. A., N. A. J. Graham, D. A. Feary, T. Morove & J. E. Cinner. 2011. Fear of fishers: human predation explains behavioral changes in coral reef fishes. PloS ONE, 6: e22761.). Furthermore, within shoals individuals are also benefited from social learning, allowing more rapid and efficient responses to a known or novel predator (Magurran, 1990Magurran, A. E. 1990. The adaptive significance of schooling as an anti-predator defence in fish. Annales Zoologici Fennici, 27: 51-66.; Hoare & Krause, 2003Hoare, D. J. & J. Krause. 2003. Social organisation, shoal structure and information transfer. Fish and Fisheries, 4: 269-279.). The communication network of behaviour is propagated by neighbours in groups and has been revealed as a process of complex information transfer (Rosenthal et al., 2015Rosenthal, S. B., C. R. Twomey, A. T. Hartnett, H. S. Wu & I. D. Couzin. 2015. Revealing the hidden networks of interaction in mobile animal groups allows prediction of complex behavioral contagion. Proceedings of the National Academy of Sciences, 112: 4690-4695.). The sharing of acquired information within the groups could be facilitated by conspecific members, although mixed groups could also increase information diversity (Sakai & Kohda, 1995Sakai, Y. & M. Kohda. 1995. Foraging by mixed-species groups involving a small Angelfish, Centropyge ferrugatus (Pomacanthidae). Japonese Journal of Ichthyology, 41: 429-435.; Hoare & Krause, 2003Hoare, D. J. & J. Krause. 2003. Social organisation, shoal structure and information transfer. Fish and Fisheries, 4: 269-279.).
Heterospecific associations have been reported in surgeonfishes, and the major functionality of such groups could be enhanced feeding and decreased predation levels (Barlow, 1974Barlow, G. W. 1974. Extraspecific imposition of social grouping among surgeonfishes (Pisces: Acanthuridae). Journal of Zoology, London, 174: 333-340.; Alevizon, 1976Alevizon, W. S. 1976. Mixed schooling and its possible significance in a tropical western Atlantic parrotfish and surgeonfish. Copeia, 1976(4): 796-798.; Itzkowitz, 1977Itzkowitz, M. 1977. Social dynamics of mixed-species groups of Jamaican reef fishes. Behavioral Ecology and Sociobiology, 2: 361-384.; Reinthal & Lewis, 1986Reinthal, P. N. & S. M. Lewis. 1986. Social behaviour, foraging efficiency and habitat utilization in a group of tropical herbivorous fish. Animal Behaviour, 34: 1687-1693.; Baird, 1993Baird, T. A. 1993. A new heterospecific foraging association between the puddingwife wrasse, Halichoeres radiatus , and the bar jack, Caranx ruber: evaluation of the foraging consequences. Environmental Biology of Fishes, 38: 393-397.). Nonetheless, there is a shortage of recent studies that strengthen the advantages for fish escape behaviour when in mixed groups (Semeniuk & Dill, 2006Semeniuk, C. A. D. & L. M. Dill. 2006. Anti-predator benefits of mixed-species groups of cowtail stingrays (Pastinachus sephen ) and whiprays (Himantura uarnak ) at rest. Ethology, 112: 33-43.). Additionally, the effect of the distance from a possible refuge on FID is reported by Gotanda et al. (2009Gotanda, K. M., K. Turgeon & D. L. Kramer. 2009. Body size and reserve protection affect flight initiation distance in parrotfishes. Behavioral Ecology and Sociobiology, 63: 1563-1572.) and it is expected to be lower when the prey is closer to a shelter (e.g. rocky crevice). In this context, habitats with greater structural complexity (i.e. presence of rocky and coral reefs) can act distinctively on antipredator behaviours and the responses may vary among species (Nunes et al., 2015Nunes, J. A. C. C., C. L. S. Sampaio & F. Barros. 2015. The influence of structural complexity and reef habitat types on flight initiation distance and escape behaviors in labrid fishes. Marine Biology, 162: 493-499.).
Some authors (e.g. Blumstein, 2006Blumstein, D. T. 2006. Developing an evolutionary ecology of fear: how life history and natural history traits affect disturbance tolerance in birds. Animal Behaviour, 71: 389-399.; Stankowich & Coss, 2007Stankowich, T. & R. G. Coss. 2007. Effects of risk assessment, predator behavior, and habitat on escape behavior in Columbian black-tailed deer. Behavioral Ecology, 18: 358-367.) consider that animals frequently respond to anthropogenic disturbance stimuli in the same way they do for natural predators, i.e. that preys follow the same economic principles they use when encountering natural predators (Frid & Dill, 2002Frid, A. & L. M. Dill. 2002. Human-caused disturbance stimuli as a form of predation risk. Conservation Ecology, 6: 11. [online]. Available from: http://www.consecol.org/vol6/iss1/art11/.
http://www.consecol.org/vol6/iss1/art11/...
). Equally, the study of Januchowski-Hartley et al. (2011Januchowski-Hartley, F. A., N. A. J. Graham, D. A. Feary, T. Morove & J. E. Cinner. 2011. Fear of fishers: human predation explains behavioral changes in coral reef fishes. PloS ONE, 6: e22761.) has highlighted some changes on fish flight behaviours, with higher FID positively associated with fishing pressure in families that are specially targeted by spear guns.
Spearfishing activity has been linked to have ecological consequences in reef fish depletion, considering abundance and fish size (Meyer, 2007Meyer, C. G. 2007. The impacts of spear and other recreational fishers on a small permanent Marine Protected Area and adjacent pulse fished area. Fisheries Research, 84: 301-307.; Frisch et al., 2008Frisch, A. J., A. J. Cole, J-P. A. Hobbs & L. Nankervis. 2008. A quantitative comparison of recreational spearfishing and linefishing on the Great Barrier Reef: implications for management of multi-sector coral reef fisheries. Coral Reefs, 27: 85-95.) and the shift on top-down control from large carnivorous toward herbivorous and omnivorous species (Godoy et al., 2010Godoy, N., S. Gelcich, J. A. Vasquez & J. C. Castilla. 2010. Spearfishing to depletion : evidence from temperate reef fishes in Chile. Ecological Applications, 20: 1504-1511.). Nevertheless, evaluating the magnitude of changes caused by this activity in marine ecosystems represents a challenge, greatly due the lack of specific management policies to regulate fishing methods (Frisch et al ., 2008Frisch, A. J., A. J. Cole, J-P. A. Hobbs & L. Nankervis. 2008. A quantitative comparison of recreational spearfishing and linefishing on the Great Barrier Reef: implications for management of multi-sector coral reef fisheries. Coral Reefs, 27: 85-95.). In this context, the knowledge of fish flight behaviours can have important implications for fishing sustainability and regulation practices, since fishes may show distinct approach distances (i.e . lower or higher FIDs) depending on target species, protection status in fished areas or the fishing intensity (Januchowski-Hartley et al., 2012Januchowski-Hartley, F. A., K. L. Nash & R. J. Lawton. 2012. Influence of spear guns, dive gear and observers on estimating fish flight initiation distance on coral reefs. Marine Ecology Progress Series, 469: 113-119.).
Here we aim to investigate the flight behaviour of a tropical reef fish, the barber surgeonfish Acanthurus bahianus Castelnau, 1855, in the presence of a spear gun diver on a popular fishing site in Northeast Brazil. We tested whether FID would be positively correlated with: (i) larger individuals; (ii) higher number of conspecifics (i.e. highest percentage of individuals of A. bahianus ) and smaller groups; (iii) with distance from a potential shelter and (iv) substrate type.
Material and Methods
Study species. The Acanthuridae are distributed in tropical and subtropical seas worldwide and comprises six genera and 80 species of Roving Herbivorous reef fishes (RHs), commonly known as Surgeonfishes (Nelson, 2006Nelson, J. S. 2006. Fishes of the world. 4th ed. Hoboken, New Jersey, John Wiley, 601p.). These roving herbivorous fishes feed primarily on filamentous benthic algae and detritus (Dias et al., 2001Dias, T. L. P., I. L. Rosa & B. M. Feitoza. 2001. Food resource and habitat sharing by the three western South Atlantic surgeonfishes (Teleostei: Acanthuridae: Acanthurus ) off Paraiba Coast, North-eastern Brazil. Aqua-Journal of Ichthyology & Aquatic Biology, 5: 1-10.; Francini-Filho et al., 2010Francini-Filho, R. B., C. M. Ferreira, E. O. C. Coni, R. L. Moura & L. Kaufman. 2010. Foraging activity of roving herbivorous reef fish (Acanthuridae and Scaridae) in eastern Brazil: influence of resource availability and interference competition. Journal of the Marine Biological Association of the United Kingdom, 90: 481-492.) and exhibit wide interspecific variation in general ecology and social behaviour (Risk, 1998Risk, A. 1998. The effects of interactions with reef residents on the settlement and subsequent persistence of ocean surgeonfish, Acanthurus bahianus . Environmental Biology of Fishes, 51: 377-389.; Alwany et al., 2005Alwany, M., E. Thaler & M. Stachowitsch. 2005. Territorial behaviour of Acanthurus sohal and Plectroglyphidodon leucozona on the fringing Egyptian red sea reefs. Environmental Biology of Fishes, 72: 321-334.).
The barber surgeonfish, Acanthurus bahianus (Acanthuridae) was the target species studied. It is found in tropical and subtropical waters of the Brazilian coast, from Parcel Manuel Luiz to Santa Catarina, including the Oceanic Islands of Fernando de Noronha, Atol das Rocas and Trindade, offshore Brazil, and Ascension and St. Helena, in the South and Central Atlantic (Sampaio & Nottingham, 2008Sampaio, C. L. S. & M. C. Nottingham. 2008. Guia para identificação de peixes ornamentais brasileiros: espécies marinhas. Brasília, DF, IBAMA, v.1 (205p).; Bernal & Rocha, 2011Bernal, M. A. & L. A. Rocha. 2011. Acanthurus tractus Poey, 1860, a valid western Atlantic species of surgeonfish (Teleostei, Acanthuridae), distinct from Acanthurus bahianus Castelnau, 1855. Zootaxa, 2905: 63-68.).
Juveniles and adults of A. bahianus are locally abundant in shallow waters (Dias et al., 2001Dias, T. L. P., I. L. Rosa & B. M. Feitoza. 2001. Food resource and habitat sharing by the three western South Atlantic surgeonfishes (Teleostei: Acanthuridae: Acanthurus ) off Paraiba Coast, North-eastern Brazil. Aqua-Journal of Ichthyology & Aquatic Biology, 5: 1-10.; Rocha et al., 2002Rocha, L. A., A. L. Bass, D. R. Robertson & B. W. Bowen, 2002. Adult habitat preferences, larval dispersal, and the comparative phylogeography of three Atlantic surgeonfishes (Teleostei: Acanthuridae). Molecular Ecology, 11: 243-252.; Francini-Filho et al., 2010Francini-Filho, R. B., C. M. Ferreira, E. O. C. Coni, R. L. Moura & L. Kaufman. 2010. Foraging activity of roving herbivorous reef fish (Acanthuridae and Scaridae) in eastern Brazil: influence of resource availability and interference competition. Journal of the Marine Biological Association of the United Kingdom, 90: 481-492.) and commonly targeted by spearfishers on south Atlantic coral reefs. Moreover, surgeonfish are known to exhibit behavioural changes as a result of fishing (Smith et al., 1989Smith, M. P. L., J. D. Bell, D. A. Pollard & B.C. Russel. 1989. Catch and effort of competition spearfishermen in Southeastern Australia. Fisheries Research, 8: 45-61.; Januchowski-Hartley et al., 2011Januchowski-Hartley, F. A., N. A. J. Graham, D. A. Feary, T. Morove & J. E. Cinner. 2011. Fear of fishers: human predation explains behavioral changes in coral reef fishes. PloS ONE, 6: e22761., 2013Januchowski-Hartley, F. A., N. A. J. Graham, J. E. Cinner& G. R. Russ. 2013. Spillover of fish naïveté from marine reserves. Ecology Letters, 16: 191-197., 2014Januchowski-Hartley, F. A., J. E. Cinner & N. A. J. Graham. 2014. Fishery benefits from behavioural modification of fishes in periodically harvested fisheries closures. Aquatic Conservation: Marine and Freshwater Ecosystems, 24: 777-710.).
Although not yet considered as fishery target species in some sites on the east coast of Brazil (Floeter et al., 2006Floeter, S. R., B. S. Halpern & C. E. L. Ferreira. 2006. Effects of fishing and protection on Brazilian reef fishes. Biological Conservation, 128: 391-402.), Acanthurus spp. are caught by inexperienced spearfishers (Bender et al., 2014Bender, M. G., G. R. Machado, P. J. A. Silva, S. R. Floeter, C. Monteiro-Netto, O. J. Luiz & C. E. L. Ferreira. 2014. Local ecological knowledge and scientific data reveal overexploitation by multigear artisanal fisheries in the Southwestern Atlantic. PLoS ONE, 9: e110332) and exported to international markets from northeast Brazil (Lessa & Nóbrega, 2000Lessa, R. & M. F. Nóbrega. 2000. Guia de identificação de peixes marinhos da Região Nordeste. Recife, Programa REVIZEE Score-NE, 128p.). Between 1996 and 2008, 3,335.49 metric tons of fish were caught and freshly exported for human consumption in the State of Rio Grande do Norte, northeastern Brazil, to the United States and the European Community, where Acanthurus spp. represented 8.64% of the total (Cunha et al., 2012Cunha, F. E. A., R. A. A. Carvalho & M. E. Araújo. 2012. Exportation of reef fish for human consumption: long-term analysis using data from Rio Grande do Norte, Brazil. Boletim do Instituto de Pesca, 38: 369-378.).
Study site. We measured Flight Initiation Distance (FID) of Acanthurus bahianus during four days in October 2014, totaling 16 hours of sampling effort, at randomly selected points distributed alongthe rocky reefs of Porto da Barra (13°23'S 38°55'W), in southeast of Todos os Santos Bay (TSB), in Salvador, Bahia, Brazil (Fig. 1). Spear gun fishing is popular in this area (Nunes et al., 2012Nunes, J. A. C. C., D. V. Medeiros, J. A. Reis-Filho, C. L. S. Sampaio & F. Barros. 2012. Reef fishes captured by recreational spearfishing on reefs of Bahia State, northeast Brazil. Biota Neotropica, 12: 179-185.) being also the focus of several reef fish studies (Coni et al., 2007Coni, E. O. C., J. A. C. C. Nunes & C. L. S. Sampaio. 2007. Halichoeres penrosei (Labridae), a sporadic cleaner wrasse. Marine Biodiversity Records, 2007: 1-2.; Nunes etal., 2007aNunes, J. A. C. C., L. C. T. Chaves, R. Maia-Nogueira & C. L. S. Sampaio. 2007a. Association between juvenile reef fish and the Caribbean reef squid Sepioteuthis sepioidea on north-eastern Brazilian coastal reefs. Journal of the Marine Biological Association of the United Kingdom, 87: 761-762., 2007bNunes, J. A. C. C., R. Maia-Nogueira & C. L. S. Sampaio. 2007b. "Friendly behaviour" between two species of Myrichthys in Brazilian waters. Coral Reefs, 26: 199.; Campos et al., 2010Campos, R. F., J. A. C. C. Nunes, D. V. Medeiros & C. L. S. Sampaio. 2010. Simultaneous reproductive event of two species of the genus Labrisomus (Labrisomidae) in Northeastern Brazil. Pan-American Journal of Aquatic Sciences, 5: 495-500.; Nunes et al ., 2013Nunes, J. A. C. C., C. L. S. Sampaio & F. Barros. 2013. How wave exposure, group size and habitat complexity influence foraging and population densities in fishes of the genus Halichoeres (Perciformes: Labridae) on tropical rocky shores. Marine Biology, 160: 2383-2394.) due to the easy access, high species richness, and shallow and clear waters.

Underwater surveys were performed at a maximum of 4 m depth and 5 m visibility. Rocky reefs are the main physiographic structure in the study sites. These habitats are covered by granite boulders and characterized by a high density of holes (Ferreira et al., 2001Ferreira, C. E. L., J. E. A. Gonçalves & R. Coutinho. 2001. Community structure of fishes and habitat complexity on a tropical rocky shore. Environmental Biology of Fishes, 61: 353-369.). Nonetheless, they represent a less complex environment than coral reefs, which are composed by builder organisms forming a tridimensional complexity reef (Nunes et al., 2015Nunes, J. A. C. C., C. L. S. Sampaio & F. Barros. 2015. The influence of structural complexity and reef habitat types on flight initiation distance and escape behaviors in labrid fishes. Marine Biology, 162: 493-499.). In this area, rocky reef substrates are predominantly composed of turf, soft coral, sea urchins, zoanthids, sponges (Nunes et al ., 2013Nunes, J. A. C. C., C. L. S. Sampaio & F. Barros. 2013. How wave exposure, group size and habitat complexity influence foraging and population densities in fishes of the genus Halichoeres (Perciformes: Labridae) on tropical rocky shores. Marine Biology, 160: 2383-2394.), small colonies of scleractinian corals and sand bottom.
Data collection. Two snorkeling divers conducted the underwater surveys. Prior to FID measurement, we estimated the total length (TL) (cm) of each target individual and the number of fish in each group (max. radius 1 m distance).
To study the heterogeneity and preferences of other species to form groups with A. bahianus , we identified the species present in the group and estimated the density of the referred species at the study site. For this, we performed 34 stationary visual censuses (adapted from Bohnsack & Bannerot, 1986Bohnsack, J. A. & S. P. Bannerot. 1986. A stationary visual census technique for quantitatively assessing community structure of coral reef fishes. NOAA Technical Report NMFS, 41: 1-15.) with 4 m radius and 5 min duration (after Nunes et al., 2013Nunes, J. A. C. C., C. L. S. Sampaio & F. Barros. 2013. How wave exposure, group size and habitat complexity influence foraging and population densities in fishes of the genus Halichoeres (Perciformes: Labridae) on tropical rocky shores. Marine Biology, 160: 2383-2394.).
The distance (cm) to the nearest potential shelter was measured as the distance between the location which the target fish started to flee from the observer's approach, and the closest ledge, hole, or vertical structure capable of providing visual isolation (Gotanda et al., 2009Gotanda, K. M., K. Turgeon & D. L. Kramer. 2009. Body size and reserve protection affect flight initiation distance in parrotfishes. Behavioral Ecology and Sociobiology, 63: 1563-1572.). Furthermore, each type of substrate (sand, turf, soft coral, sea urchin, zoanthids or sponges) at the site of observation (max. radius 1 m distance) had its coverage percentage visually estimated.
Flight initiation distance. The same diver measured the FID of A. bahianus individuals. A potential target fish was first identified from the surface. A diver would then descend to the same depth as the targeted fish and swim towards the fish at a constant speed (Januchowski-Hartley et al., 2011Januchowski-Hartley, F. A., N. A. J. Graham, D. A. Feary, T. Morove & J. E. Cinner. 2011. Fear of fishers: human predation explains behavioral changes in coral reef fishes. PloS ONE, 6: e22761., 2012Januchowski-Hartley, F. A., K. L. Nash & R. J. Lawton. 2012. Influence of spear guns, dive gear and observers on estimating fish flight initiation distance on coral reefs. Marine Ecology Progress Series, 469: 113-119.). The diver was dressed in a spearfishing wetsuit and reproduced spearfisher behavior, remaining approximately 3 m far, holding the spear gun horizontally in front of the face and pointing directly to the targeted fish. When the fish fled, the diver placed a marker on the substrate directly below the point where their hand was holding the gun at the moment of flight, and then a second marker was left on the substrate directly below the location of the fish when it fled (Januchowski-Hartley et al ., 2011Januchowski-Hartley, F. A., N. A. J. Graham, D. A. Feary, T. Morove & J. E. Cinner. 2011. Fear of fishers: human predation explains behavioral changes in coral reef fishes. PloS ONE, 6: e22761., 2012Januchowski-Hartley, F. A., K. L. Nash & R. J. Lawton. 2012. Influence of spear guns, dive gear and observers on estimating fish flight initiation distance on coral reefs. Marine Ecology Progress Series, 469: 113-119.). The distance (cm) between these markers was measured using a graduated tape.
The spear gun used in the survey was 46 cm and this distance was therefore subtracted from the measurements to obtain the FID. To avoid recording fishes from the same group in quick succession, the diver immediately moved at least 5 m away after each observation (Nunes et al., 2013Nunes, J. A. C. C., C. L. S. Sampaio & F. Barros. 2013. How wave exposure, group size and habitat complexity influence foraging and population densities in fishes of the genus Halichoeres (Perciformes: Labridae) on tropical rocky shores. Marine Biology, 160: 2383-2394.).
Data analysis. FID data was tested for normality through Shapiro-Wilk W test (W = 0.9296; p = 0.08) and homogeneity of variance was determined using Levene's test (F (1.72) = 0.293; p = 0.589). Analysis of covariance (ANCOVA) was used to investigate three continuous covariates: the influence of size of target fish (TL), group size and distance of shelter, and one categorical variable: substrate type, on FID of A. bahianus . Tukey's test was used to identify differences in FID.
We used a T-test to compare the FID between interspecific and mono specific shoals. One-way analysis of variance (ANOVA) was performed to investigate whether there were differences in FID in different levels of specificity of groups in the absence of Acanthurus bahianus , i.e. the percentage of individuals of other species in each group (0-30%, 31-60%, > 60%). Additionally, an electivity index, Ivlev's, was used to identify the group formation preferences.
Ivlev's was calculated using the formula: E i = (ri - ni )/(ri + ni), where E i is the value of electivity for the selection of species i in group formation with A. bahianus ; r i is the percentage of encounters of species i with A. bahianus and n i is the relative density of species i in area. Electivity index varied from -1 to 1. Values close to -1 show low preference, or rejection, while values close to 1 indicate high preference for a group formation with A. bahianus (Krebs, 1989Krebs, C. J. 1989. Ecological methodology. New York, Harper Collins, 654p.). T-test and ANCOVA were performed in StatSoft STATISTICA, version 8.0, and all data analyses were performed considering a significance level α ≤ 0.05.
Results
Body size, group size and distance to shelter. We measured the FID of 74 individuals of A. bahianus , which ranged from 6 to 30 cm TL and were solitary or present in groups ranging from 2 to 34 individuals. The results of Analysis of covariance (Table 1) showed significant differences in FID between some of the variables (p < 0.001). We found a significant and positive correlation between FID and body size, with an increase in FID in large-bodied individuals (Fig. 2b).
There was no significant difference between FID and group size or in distance to a potential shelter (Table 1). However, FID varied between the main substrates (ANCOVA; F = 3.95, p = 0.028) (Table 1) with greater flight distances observed where there was a higher percentage of sand substrate (Fig. 3).
Analysis of covariance (ANCOVA) results testing the influence of fish size, group size, distance of shelter and substrate on Flight Initiation Distance of Acanthurus bahianus . SS = Sum of squares; Df = Degree of freedom; MS = Mean of squares. *Significant difference.

Flight Initiation Distance (cm) plotted against Group size (a), Body size (cm) (b), and Distance of shelter (cm) (c) for Acanthurus bahianus .

Flight Initiation Distance (cm) (mean ± S.D.) for Acanthurus bahianus in the main substrates found at the study site. Different letters means significant differences - p < 0.05.
Composition and preference in a group formation. The species composition in groups did not significantly affect the FID of A. bahianus (One-way ANOVA; F = 0.097, p = 0.908) based on three categories of mixed species groups (mean ± S.D.): 0-30% Low (39.4 ± 28.1); 31-60% Medium (37.5 ± 27.3); > 60% High (36.6 ± 23.5). Furthermore, there were no differences between FID of interspecific and mono specific shoals (t = 0.0499; df = 66.00; p = 0.619).
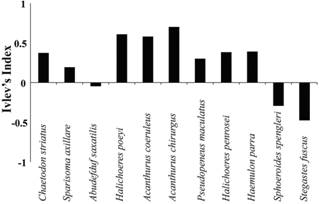
On the other hand, Ivlev's electivity index showed that 3 out of 11 species had a strong preference for group formation with A. bahianus: Acanthurus chirurgus , Halichoeres poeyi and Acanthurus coeruleus , in order of preference, respectively.Two species showed no preferences: Stegastes fuscus and Sphoeroides spengleri (Fig. 4).

Results of Ivlev's index showing the preference of Acanthurus chirurgus, Acanthurus coeruleus and Halichoeres poey ; and the rejection of Stegastes fuscus and Sphoeroides spengleri to form groups with Acanthurus bahianus .
Discussion
We investigated FID of Acanthurus bahianus when confronted with a spearfisher. Our results indicate that: (i) body size significantly influence FID, with smaller individuals having lower FID than larger ones; (ii) size and group composition did not affect the escape response of A. bahianus , albeit congeneric species of Acanthurus , and H. poeyi were seen in groups with A. bahianus ; and (iii) despite the distance to shelter having no influence on FID, escape responses varied between main substrate types (Table 1).
Greater FID was often associated with larger body size (Fig. 2b), which could be attributed to spearfisher preferences in targeting larger fishes (Januchowski-hartley et al., 2011Januchowski-Hartley, F. A., N. A. J. Graham, D. A. Feary, T. Morove & J. E. Cinner. 2011. Fear of fishers: human predation explains behavioral changes in coral reef fishes. PloS ONE, 6: e22761.), however our observations may also be alternatively explained. Large-bodied species can be correlated with age at maturity or age at the first reproduction and these life history traits could affect the fearfulness, making species reduce the risk-taking (Blumstein, 2006Blumstein, D. T. 2006. Developing an evolutionary ecology of fear: how life history and natural history traits affect disturbance tolerance in birds. Animal Behaviour, 71: 389-399.). In the same way, the reproductive value strongly supports explaining the increase in FID with body size in parrotfishes (Gotanda et al., 2009Gotanda, K. M., K. Turgeon & D. L. Kramer. 2009. Body size and reserve protection affect flight initiation distance in parrotfishes. Behavioral Ecology and Sociobiology, 63: 1563-1572.). Furthemore, experience or social learning related to predator attacks may provide an opportunity for learnt escape responses in adult fish (Kelley & Magurran, 2003Kelley, J. L. & A. E. Magurran. 2003. Learned predator recognition and antipredator responses in fishes. Fish and Fisheries, 4: 216-226.).
Although a relationship between FID and shoal size has not been found here, previous studies have shown that fishes can perform coordinated shoaling behaviour on different shoal sizes in response to increased predation risk (Magurran & Pitcher, 1987Magurran, A. E. & T. J. Pitcher. 1987. Provenance, shoal size and the sociobiology of predator-evasion behaviour in minnow shoals. Proceedings of the Royal Society of London. Series B: Biological Sciences, 229: 439-465.). One possible explanation for this group effect is the 'many-eyes hypothesis' (Pulliam, 1973Pulliam, H. R. 1973. On the advantages of flocking. Journal of Theoretical Biology, 38: 419-422.), which states that as the group size increases, there are more eyes scanning the environment for predators (Lima, 1995Lima, S. L. 1995. Back to the basics of anti-predatory vigilance: the group-size effect. Animal Behaviour, 49: 11-20.). Thus, the earlier collective detection of a predator attack and the individual perception of safety in groups (Magurran, 1990Magurran, A. E. 1990. The adaptive significance of schooling as an anti-predator defence in fish. Annales Zoologici Fennici, 27: 51-66.) may result in a lower FID. In conformity with this prediction, Wolf (1987Wolf, N. G. 1987. Schooling tendency and foraging benefit in the ocean surgeonfish. Behavioral Ecology and Sociobiology, 21: 59-63.) reports that juveniles of a sister species of barber surgeonfish, Acanthurus tractus (see Bernal & Rocha, 2011Bernal, M. A. & L. A. Rocha. 2011. Acanthurus tractus Poey, 1860, a valid western Atlantic species of surgeonfish (Teleostei, Acanthuridae), distinct from Acanthurus bahianus Castelnau, 1855. Zootaxa, 2905: 63-68.) can benefit from having more time to forage instead of vigilance when in schooling groups than solitary ones.
Nevertheless, the relationship between group size and escape distance is controversial and may generate contrasting results due to variables related to shoaling (e.g. variables affecting vigilance) (Roberts, 1996Roberts, G. 1996. Why individual vigilance declines as group size increases. Animal Behaviour, 51: 1077-1086.). For instance, Semeniuk & Dill (2005Semeniuk, C. A. D. & L. M. Dill. 2005. Cost/benefit analysis of group and solitary resting in the cowtail stingray, Pastinachus sephen . Behavioral Ecology, 16: 417-426.) found longer reaction distances in groups than in solitary individuals of cowtail stingray, Pastinachus sephen . Similarly, Januchowski-Hartley et al. (2011Januchowski-Hartley, F. A., N. A. J. Graham, D. A. Feary, T. Morove & J. E. Cinner. 2011. Fear of fishers: human predation explains behavioral changes in coral reef fishes. PloS ONE, 6: e22761.) observed an increase of FID with growth on group size for Acanthuridae. The latter study suggested that this result could indicate an independent anti-predation response to variation in fishing pressure.
Living in conspecific or heterospecific aggregations can facilitate social learning of antipredator behaviour through chemical and visual cues (Griffin, 2004Griffin, A. S. 2004. Social learning about predators: a review and prospectus. Learning & Behavior, 32: 131-140.; Lönnstedt & McCormick, 2011Lönnstedt, O. M. & M. I. McCormick. 2011. Chemical alarm cues inform prey of predation threat: the importance of ontogeny and concentration in a coral reef fish. Animal Behaviour, 82: 213-218.). Monospecific grouping may provide foraging and anti-predator benefits, however mixed-species groups are probably more advantageous, since they create less resource competition and provide significant benefits such as confusing predators by preventing them to focus on a particular individual, an earlier detection of potential threats and greater flight distances to predators (Semeniuk & Dill, 2005Semeniuk, C. A. D. & L. M. Dill. 2005. Cost/benefit analysis of group and solitary resting in the cowtail stingray, Pastinachus sephen . Behavioral Ecology, 16: 417-426., 2006Semeniuk, C. A. D. & L. M. Dill. 2006. Anti-predator benefits of mixed-species groups of cowtail stingrays (Pastinachus sephen ) and whiprays (Himantura uarnak ) at rest. Ethology, 112: 33-43.). To illustrate, studies of heterospecific schooling have indicated that associations of Rusty Angelfish, Centropyge ferrugatus , with mixed-species groups provide an antipredatory and foraging benefit (Sakai & Kohda, 1995Sakai, Y. & M. Kohda. 1995. Foraging by mixed-species groups involving a small Angelfish, Centropyge ferrugatus (Pomacanthidae). Japonese Journal of Ichthyology, 41: 429-435.).
Assuming that species can learn that spearfishers represent a potential threat, it is possible that the preference of species found here to form groups with barber surgeonfish (Fig. 4) could mutually reinforce appropriate antipredator responses. Barber surgeonfishes are often accompanied by A. chirurgus and A. coeruleus (Dias et al., 2001Dias, T. L. P., I. L. Rosa & B. M. Feitoza. 2001. Food resource and habitat sharing by the three western South Atlantic surgeonfishes (Teleostei: Acanthuridae: Acanthurus ) off Paraiba Coast, North-eastern Brazil. Aqua-Journal of Ichthyology & Aquatic Biology, 5: 1-10.) and the relative frequencies of each species in a group can vary with size classes. This allows mixed groups to exploit territories with high resource quality (i.e. higher biomass and algal diversity) (Wolf, 1987Wolf, N. G. 1987. Schooling tendency and foraging benefit in the ocean surgeonfish. Behavioral Ecology and Sociobiology, 21: 59-63.; Ferreira et al., 1998Ferreira, C. E. L., J. E. A. Gonçalves, R. Coutinho & A. C. Peret. 1998. Herbivory by the Dusky Damselfish Stegastes fuscus (Cuvier, 1830) in a tropical rocky shore: effects on the benthic community. Journal of Experimental Marine Biology and Ecology, 229: 241-264.). Similarly, the willingness of Halichoeres species to form groups with roving herbivores, such as Surgeonfish is well known, and can be explained by the associated decrease in predation risk and increase in foraging opportunities (Alevizon, 1976Alevizon, W. S. 1976. Mixed schooling and its possible significance in a tropical western Atlantic parrotfish and surgeonfish. Copeia, 1976(4): 796-798.; Dias et al ., 2001Dias, T. L. P., I. L. Rosa & B. M. Feitoza. 2001. Food resource and habitat sharing by the three western South Atlantic surgeonfishes (Teleostei: Acanthuridae: Acanthurus ) off Paraiba Coast, North-eastern Brazil. Aqua-Journal of Ichthyology & Aquatic Biology, 5: 1-10.; Nunes et al., 2013Nunes, J. A. C. C., C. L. S. Sampaio & F. Barros. 2013. How wave exposure, group size and habitat complexity influence foraging and population densities in fishes of the genus Halichoeres (Perciformes: Labridae) on tropical rocky shores. Marine Biology, 160: 2383-2394.). Furthermore, Halichoeres spp. are often observed (including on our study site) engaged in cleaning activities with regular client fish such as A.bahianus (Sazima et al., 1998Sazima, I., R. L. Moura& J. L. Gasparini. 1998. The wrasse Halichoeres cyanocephalus (Labridae) as a specialized cleaner fish. Bulletin of Marine Science, 63: 605-610.; Coni et al., 2007Coni, E. O. C., J. A. C. C. Nunes & C. L. S. Sampaio. 2007. Halichoeres penrosei (Labridae), a sporadic cleaner wrasse. Marine Biodiversity Records, 2007: 1-2.).
Groups of barber surgeonfish can consume large amounts of algal biomass from damselfish (Stegastes fuscus ) territories (Lawson et al., 1999Lawson, G. L., D. L. Kramer & W. Hunte. 1999. Size-related habitat use and schooling behavior in two species of surgeonfish (Acanthurus bahianus and A. coeruleus ) on a fringing reef in Barbados, West Indies. Environmental Biology of Fishes, 54: 19-33.; Dias et al., 2001Dias, T. L. P., I. L. Rosa & B. M. Feitoza. 2001. Food resource and habitat sharing by the three western South Atlantic surgeonfishes (Teleostei: Acanthuridae: Acanthurus ) off Paraiba Coast, North-eastern Brazil. Aqua-Journal of Ichthyology & Aquatic Biology, 5: 1-10.). This antagonistic behaviour explains the lack of association between S. fuscus and A. bahianus (Fig. 4), despite the very high abundance of Brazilian damselfish (Ferreira et al., 1998Ferreira, C. E. L., J. E. A. Gonçalves, R. Coutinho & A. C. Peret. 1998. Herbivory by the Dusky Damselfish Stegastes fuscus (Cuvier, 1830) in a tropical rocky shore: effects on the benthic community. Journal of Experimental Marine Biology and Ecology, 229: 241-264.; Osório et al., 2006Osório, R., I. L. Rosa& H. Cabral. 2006. Territorial defence by the Brazilian damsel Stegastes fuscus (Teleostei: Pomacentridae). Journal of Fish Biology, 69: 233-242.). The Bandtail puffer, Sphoeroides spengleri , a common local species that feeds opportunistically on fish (Randall, 1967Randall, J. E. 1967. Food habits of reef fishes of the West Indies. Studies in Tropical Oceanography, 5: 665-847.) is also avoided, possibly due the risk to be bite or other antagonistic behavior.
To have refuge into crevices or undersurface of boulders is apparently effective for some species (e.g. sea breams) in order to combat the attacks by large predators (Guidetti et al., 2008Guidetti, P., E. Vierucci & S. Bussotti. 2008. Differences in escape response of fish in protected and fished Mediterranean rocky reefs. Journal of the Marine Biological Association of the United Kingdom, 88: 625-627.). The assumption that flight initiation distance is associated with structural complexity is supported by other studies, where Labridae fish showed a decrease on FID in structurally higher habitats (Nunes et al., 2015Nunes, J. A. C. C., C. L. S. Sampaio & F. Barros. 2015. The influence of structural complexity and reef habitat types on flight initiation distance and escape behaviors in labrid fishes. Marine Biology, 162: 493-499.), however the relationship between FID with distance to rocky reefs for A. bahianus was not significant(Table 1; Fig. 2c). The natural structure and physical complexity of rocky reefs, mainly determined by their density of holes, are less complex than coral reefs are (Ferreira et al., 2001Ferreira, C. E. L., J. E. A. Gonçalves & R. Coutinho. 2001. Community structure of fishes and habitat complexity on a tropical rocky shore. Environmental Biology of Fishes, 61: 353-369.; Floeter et al., 2004Floeter, S. R., C. E. L. Ferreira, A. Dominici-Arosemena & I. R. Zalmon. 2004. Latitudinal gradients in Atlantic reef fish communities: trophic structure and spatial use patterns. Journal of Fish Biology, 64: 1680-1699.) and it is possible that A. bahianus does not consider the available crevices and overhangs as potential shelters capable of providing protection from predation.
In contrast, we found higher FID associated with more homogenous substrates (i.e. sand bottom) (Fig. 3). Possibly, because no shelter was available in these points, A. bahianus preferred avoid taking the risk of a close approach from the spearfisher and escaped as soon as the threat was detected.
Acanthurids are abundant and important members of the reef community, playing significant ecological roles in coral reef resilience. They graze epilithic algal turfs and limit the establishment and growth of macroalgae that can kill coral colonies by preventing recruitment (Green & Bellwood, 2009Green, A. L. & D. R. Bellwood. 2009. Monitoring functional groups of herbivorous reef fishes as indicators of coral reef resilience: a practical guide for coral reef managers in the Asia Pacific region. Gland, Switzerland, Working Group on Climate Change and Coral Reefs, International Union for the Conservation of Nature (IUCN), 69p. (IUCN Resilience Science Group Working Paper Series - nº 7). Available from: http://cmsdata.iucn.org/downloads/resilience_herbivorous_monitoring.pdf.
http://cmsdata.iucn.org/downloads/resili...
). Fishing for Acanthurus spp. in Brazil has been described by many authors (e.g. Ribeiro, 2004Ribeiro, F. P. 2004. Composição da biocenose e abundância relativa de peixes capturados com covos nos estados do Rio Grande do Norte e Pernambuco (Brasil). Boletim Técnico-Científico do CEPENE, 12: 113-128.; Nóbrega & Lessa, 2007Nóbrega, M. F. & R. P. Lessa. 2007. Descrição e composição das capturas da frota pesqueira artesanal da região Nordeste do Brasil. Arquivos de Ciências do Mar, 40: 64-74.; Cunha et al., 2012Cunha, F. E. A., R. A. A. Carvalho & M. E. Araújo. 2012. Exportation of reef fish for human consumption: long-term analysis using data from Rio Grande do Norte, Brazil. Boletim do Instituto de Pesca, 38: 369-378.) and goes back for more than a decade in the artisanal fisheries of states in the "Hump of Brazil". They are also under increasing threat as a result of fishing for multiple uses (Sampaio & Rosa, 2005Sampaio, C. L. S. & I. L. Rosa. 2005. A coleta de peixes ornamentais marinhos na Bahia, Brasil: técnicas utilizadas e implicações à conservação. Boletim Técnico-Científico do CEPENE, 13: 39-51.; Sampaio & Nottingham, 2008Sampaio, C. L. S. & M. C. Nottingham. 2008. Guia para identificação de peixes ornamentais brasileiros: espécies marinhas. Brasília, DF, IBAMA, v.1 (205p).; Cunha et al ., 2012Cunha, F. E. A., R. A. A. Carvalho & M. E. Araújo. 2012. Exportation of reef fish for human consumption: long-term analysis using data from Rio Grande do Norte, Brazil. Boletim do Instituto de Pesca, 38: 369-378.).
Reduction in population of medium-large herbivore species such as A. bahianus by spearfishing might result in a significant loss of ecosystem functions (Green & Bellwood, 2009Green, A. L. & D. R. Bellwood. 2009. Monitoring functional groups of herbivorous reef fishes as indicators of coral reef resilience: a practical guide for coral reef managers in the Asia Pacific region. Gland, Switzerland, Working Group on Climate Change and Coral Reefs, International Union for the Conservation of Nature (IUCN), 69p. (IUCN Resilience Science Group Working Paper Series - nº 7). Available from: http://cmsdata.iucn.org/downloads/resilience_herbivorous_monitoring.pdf.
http://cmsdata.iucn.org/downloads/resili...
; Burkepile & Hay, 2011Burkepile, D. E. & M. E. Hay. 2011. Feeding complementarity versus redundancy among herbivorous fishes on a Caribbean reef. Coral Reefs, 30: 351-362.; Bonaldo et al., 2014Bonaldo, R. M., A. S. Hoey & D. R. Bellwood. 2014. The ecosystem roles of parrotfishes on tropical reefs. Oceanography and Marine Biology: An Annual Review, 52: 81-132.). Additionally, an increase in wariness in target species such as A. bahianus may reduce fishing success and make fishers shift their preferences. This could affect antipredator behaviour of (currently) non-target species and increase their sensitivity to spearfisher disturbance.
In conclusion, our study provides evidence that A. bahianus alters their perception of predation risk with increased body size or when in areas of low physical complexity. However, the relationship between FID and group composition is not yet completely understood: FID does not seem to be influenced by group composition per se, but the preferences in group formation shows clear selectivity for living in specific shoals. We highlight that future research should focus on the fish preferences in group formation, e.g. aggregations in mixed or single species shoals, and on indirect impacts of spearfishing on the structure of marine communities, emphasizing in considering aggregation behaviours and the social learning of juvenile and adult fishes as an important tool to avoid predation.
Acknowledgements
We thank Dive Bahia team for supporting during fieldwork, and CAPES for the scholarship provided to LJB and JACCN. Thanks are extended to Gabriel C. C. Ferreira (UFES) and Taciana K. Pinto (UFAL) for help with figures design and analysis; Liliana Colman (University of Exeter, UK) for reviewing and improving English; and two anonymous reviewers that provided comments which improved this manuscript.
References
- Abrams, P. A. 2000. The evolution of predator-prey interactions: theory and evidence. Annual Review of Ecology and Systematics, 31: 79-105.
- Alevizon, W. S. 1976. Mixed schooling and its possible significance in a tropical western Atlantic parrotfish and surgeonfish. Copeia, 1976(4): 796-798.
- Alwany, M., E. Thaler & M. Stachowitsch. 2005. Territorial behaviour of Acanthurus sohal and Plectroglyphidodon leucozona on the fringing Egyptian red sea reefs. Environmental Biology of Fishes, 72: 321-334.
- Baird, T. A. 1993. A new heterospecific foraging association between the puddingwife wrasse, Halichoeres radiatus , and the bar jack, Caranx ruber: evaluation of the foraging consequences. Environmental Biology of Fishes, 38: 393-397.
- Barlow, G. W. 1974. Extraspecific imposition of social grouping among surgeonfishes (Pisces: Acanthuridae). Journal of Zoology, London, 174: 333-340.
- Bateman, P. W. & P. A. Fleming. 2014. Living on the edge: effects of body size, group density and microhabitat selection on escape behaviour of southern leopard frogs Lithobates sphenocephalus . Current Zoology, 60: 712-718.
- Bender, M. G., G. R. Machado, P. J. A. Silva, S. R. Floeter, C. Monteiro-Netto, O. J. Luiz & C. E. L. Ferreira. 2014. Local ecological knowledge and scientific data reveal overexploitation by multigear artisanal fisheries in the Southwestern Atlantic. PLoS ONE, 9: e110332
- Bernal, M. A. & L. A. Rocha. 2011. Acanthurus tractus Poey, 1860, a valid western Atlantic species of surgeonfish (Teleostei, Acanthuridae), distinct from Acanthurus bahianus Castelnau, 1855. Zootaxa, 2905: 63-68.
- Blumstein, D. T. 2006. Developing an evolutionary ecology of fear: how life history and natural history traits affect disturbance tolerance in birds. Animal Behaviour, 71: 389-399.
- Blumstein, D. T., E. Fernández-Juricic, P. A. Zollner & S. C. Garity. 2005. Inter-specific variation in avian responses to human disturbance. Journal of Applied Ecology, 42: 943-953.
- Bohnsack, J. A. & S. P. Bannerot. 1986. A stationary visual census technique for quantitatively assessing community structure of coral reef fishes. NOAA Technical Report NMFS, 41: 1-15.
- Bonaldo, R. M., A. S. Hoey & D. R. Bellwood. 2014. The ecosystem roles of parrotfishes on tropical reefs. Oceanography and Marine Biology: An Annual Review, 52: 81-132.
- Brown, J. S., B. P. Kotler & A. Bouskila. 2001. Ecology of fear: foraging games between predators and prey with pulsed resources. Annales Zoologici Fennici,38: 71-87.
- Brown, C. & K. N. Laland. 2003. Social learning in fishes: a review. Fish and Fisheries, 4: 280-288.
- Burkepile, D. E. & M. E. Hay. 2011. Feeding complementarity versus redundancy among herbivorous fishes on a Caribbean reef. Coral Reefs, 30: 351-362.
- Campos, R. F., J. A. C. C. Nunes, D. V. Medeiros & C. L. S. Sampaio. 2010. Simultaneous reproductive event of two species of the genus Labrisomus (Labrisomidae) in Northeastern Brazil. Pan-American Journal of Aquatic Sciences, 5: 495-500.
- Coni, E. O. C., J. A. C. C. Nunes & C. L. S. Sampaio. 2007. Halichoeres penrosei (Labridae), a sporadic cleaner wrasse. Marine Biodiversity Records, 2007: 1-2.
- Cooper, W. E., Jr. 2006. Dynamic risk assessment: prey rapidly adjust flight initiation distance to changes in predator approach speed. Ethology, 112: 858-864.
- Cooper, W. E., Jr. 2009. Flight initiation distance decreases during social activity in lizards (Sceloporus virgatus ). Behavioral Ecology and Sociobiology, 63: 1765-1771.
- Cooper, W. E., Jr. & W. G. Frederick. 2007. Optimal flight initiation distance. Journal of Theoretical Biology, 244: 59-67.
- Cooper, W. E., Jr. & T. Stankowich. 2010. Prey or predator? Body size of an approaching animal affects decisions to attack or escape. Behavioral Ecology, 21: 1278-1284.
- Cunha, F. E. A., R. A. A. Carvalho & M. E. Araújo. 2012. Exportation of reef fish for human consumption: long-term analysis using data from Rio Grande do Norte, Brazil. Boletim do Instituto de Pesca, 38: 369-378.
- Dias, T. L. P., I. L. Rosa & B. M. Feitoza. 2001. Food resource and habitat sharing by the three western South Atlantic surgeonfishes (Teleostei: Acanthuridae: Acanthurus ) off Paraiba Coast, North-eastern Brazil. Aqua-Journal of Ichthyology & Aquatic Biology, 5: 1-10.
- Dill, L. M. 1974. The escape response of the zebra danio (Brachydanio rerio ) II. The effect of experience. Animal Behaviour, 22: 723-730.
- Domenici, P. 2010. Context-dependent variability in the components of fish escape response: integrating locomotor performance and behavior. Journal of Experimental Zoology Part A: Ecological Genetics and Physiology, 313: 59-79.
- Domenici, P., C. Lefrançois & A. Shingles. 2007. Hypoxia and the antipredator behaviours of fishes. Philosophical Transactions of the Royal Society B: Biological Sciences, 362: 2105-2121.
- Feary, D. A., J. E. Cinner, N. A. J. Graham & F. A. Januchowski-Hartley. 2011. Effects of customary marine closures on fish behavior, spear-fishing success, and underwater visual surveys. Conservation Biology, 25: 341-349.
- Ferreira, C. E. L., J. E. A. Gonçalves, R. Coutinho & A. C. Peret. 1998. Herbivory by the Dusky Damselfish Stegastes fuscus (Cuvier, 1830) in a tropical rocky shore: effects on the benthic community. Journal of Experimental Marine Biology and Ecology, 229: 241-264.
- Ferreira, C. E. L., J. E. A. Gonçalves & R. Coutinho. 2001. Community structure of fishes and habitat complexity on a tropical rocky shore. Environmental Biology of Fishes, 61: 353-369.
- Floeter, S. R., C. E. L. Ferreira, A. Dominici-Arosemena & I. R. Zalmon. 2004. Latitudinal gradients in Atlantic reef fish communities: trophic structure and spatial use patterns. Journal of Fish Biology, 64: 1680-1699.
- Floeter, S. R., B. S. Halpern & C. E. L. Ferreira. 2006. Effects of fishing and protection on Brazilian reef fishes. Biological Conservation, 128: 391-402.
- Francini-Filho, R. B., C. M. Ferreira, E. O. C. Coni, R. L. Moura & L. Kaufman. 2010. Foraging activity of roving herbivorous reef fish (Acanthuridae and Scaridae) in eastern Brazil: influence of resource availability and interference competition. Journal of the Marine Biological Association of the United Kingdom, 90: 481-492.
- Frid, A. & L. M. Dill. 2002. Human-caused disturbance stimuli as a form of predation risk. Conservation Ecology, 6: 11. [online]. Available from: http://www.consecol.org/vol6/iss1/art11/
» http://www.consecol.org/vol6/iss1/art11/ - Frisch, A. J., A. J. Cole, J-P. A. Hobbs & L. Nankervis. 2008. A quantitative comparison of recreational spearfishing and linefishing on the Great Barrier Reef: implications for management of multi-sector coral reef fisheries. Coral Reefs, 27: 85-95.
- Godoy, N., S. Gelcich, J. A. Vasquez & J. C. Castilla. 2010. Spearfishing to depletion : evidence from temperate reef fishes in Chile. Ecological Applications, 20: 1504-1511.
- Gotanda, K. M., K. Turgeon & D. L. Kramer. 2009. Body size and reserve protection affect flight initiation distance in parrotfishes. Behavioral Ecology and Sociobiology, 63: 1563-1572.
- Green, A. L. & D. R. Bellwood. 2009. Monitoring functional groups of herbivorous reef fishes as indicators of coral reef resilience: a practical guide for coral reef managers in the Asia Pacific region. Gland, Switzerland, Working Group on Climate Change and Coral Reefs, International Union for the Conservation of Nature (IUCN), 69p. (IUCN Resilience Science Group Working Paper Series - nº 7). Available from: http://cmsdata.iucn.org/downloads/resilience_herbivorous_monitoring.pdf
» http://cmsdata.iucn.org/downloads/resilience_herbivorous_monitoring.pdf - Griffin, A. S. 2004. Social learning about predators: a review and prospectus. Learning & Behavior, 32: 131-140.
- Guidetti, P., E. Vierucci & S. Bussotti. 2008. Differences in escape response of fish in protected and fished Mediterranean rocky reefs. Journal of the Marine Biological Association of the United Kingdom, 88: 625-627.
- Hoare, D. J. & J. Krause. 2003. Social organisation, shoal structure and information transfer. Fish and Fisheries, 4: 269-279.
- Itzkowitz, M. 1977. Social dynamics of mixed-species groups of Jamaican reef fishes. Behavioral Ecology and Sociobiology, 2: 361-384.
- Januchowski-Hartley, F. A., J. E. Cinner & N. A. J. Graham. 2014. Fishery benefits from behavioural modification of fishes in periodically harvested fisheries closures. Aquatic Conservation: Marine and Freshwater Ecosystems, 24: 777-710.
- Januchowski-Hartley, F. A., N. A. J. Graham, J. E. Cinner& G. R. Russ. 2013. Spillover of fish naïveté from marine reserves. Ecology Letters, 16: 191-197.
- Januchowski-Hartley, F. A., N. A. J. Graham, D. A. Feary, T. Morove & J. E. Cinner. 2011. Fear of fishers: human predation explains behavioral changes in coral reef fishes. PloS ONE, 6: e22761.
- Januchowski-Hartley, F. A., K. L. Nash & R. J. Lawton. 2012. Influence of spear guns, dive gear and observers on estimating fish flight initiation distance on coral reefs. Marine Ecology Progress Series, 469: 113-119.
- Kelley, J. L. & A. E. Magurran. 2003. Learned predator recognition and antipredator responses in fishes. Fish and Fisheries, 4: 216-226.
- Krebs, C. J. 1989. Ecological methodology. New York, Harper Collins, 654p.
- Lawson, G. L., D. L. Kramer & W. Hunte. 1999. Size-related habitat use and schooling behavior in two species of surgeonfish (Acanthurus bahianus and A. coeruleus ) on a fringing reef in Barbados, West Indies. Environmental Biology of Fishes, 54: 19-33.
- Lessa, R. & M. F. Nóbrega. 2000. Guia de identificação de peixes marinhos da Região Nordeste. Recife, Programa REVIZEE Score-NE, 128p.
- Lima, S. L. 1995. Back to the basics of anti-predatory vigilance: the group-size effect. Animal Behaviour, 49: 11-20.
- Lönnstedt, O. M. & M. I. McCormick. 2011. Chemical alarm cues inform prey of predation threat: the importance of ontogeny and concentration in a coral reef fish. Animal Behaviour, 82: 213-218.
- Magurran, A. E. 1990. The adaptive significance of schooling as an anti-predator defence in fish. Annales Zoologici Fennici, 27: 51-66.
- Magurran, A. E. & T. J. Pitcher. 1987. Provenance, shoal size and the sociobiology of predator-evasion behaviour in minnow shoals. Proceedings of the Royal Society of London. Series B: Biological Sciences, 229: 439-465.
- Marras, S., S. S. Killen, G. Claireaux, P. Domenici & D. J. McKenzie. 2011. Behavioural and kinematic components of the fast-start escape response in fish: individual variation and temporal repeatability. Journal of Experimental Biology, 214: 3102-3110.
- Meyer, C. G. 2007. The impacts of spear and other recreational fishers on a small permanent Marine Protected Area and adjacent pulse fished area. Fisheries Research, 84: 301-307.
- Nelson, J. S. 2006. Fishes of the world. 4th ed. Hoboken, New Jersey, John Wiley, 601p.
- Nóbrega, M. F. & R. P. Lessa. 2007. Descrição e composição das capturas da frota pesqueira artesanal da região Nordeste do Brasil. Arquivos de Ciências do Mar, 40: 64-74.
- Nunes, J. A. C. C., L. C. T. Chaves, R. Maia-Nogueira & C. L. S. Sampaio. 2007a. Association between juvenile reef fish and the Caribbean reef squid Sepioteuthis sepioidea on north-eastern Brazilian coastal reefs. Journal of the Marine Biological Association of the United Kingdom, 87: 761-762.
- Nunes, J. A. C. C., R. Maia-Nogueira & C. L. S. Sampaio. 2007b. "Friendly behaviour" between two species of Myrichthys in Brazilian waters. Coral Reefs, 26: 199.
- Nunes, J. A. C. C., D. V. Medeiros, J. A. Reis-Filho, C. L. S. Sampaio & F. Barros. 2012. Reef fishes captured by recreational spearfishing on reefs of Bahia State, northeast Brazil. Biota Neotropica, 12: 179-185.
- Nunes, J. A. C. C., C. L. S. Sampaio & F. Barros. 2013. How wave exposure, group size and habitat complexity influence foraging and population densities in fishes of the genus Halichoeres (Perciformes: Labridae) on tropical rocky shores. Marine Biology, 160: 2383-2394.
- Nunes, J. A. C. C., C. L. S. Sampaio & F. Barros. 2015. The influence of structural complexity and reef habitat types on flight initiation distance and escape behaviors in labrid fishes. Marine Biology, 162: 493-499.
- Osório, R., I. L. Rosa& H. Cabral. 2006. Territorial defence by the Brazilian damsel Stegastes fuscus (Teleostei: Pomacentridae). Journal of Fish Biology, 69: 233-242.
- Pulliam, H. R. 1973. On the advantages of flocking. Journal of Theoretical Biology, 38: 419-422.
- Randall, J. E. 1967. Food habits of reef fishes of the West Indies. Studies in Tropical Oceanography, 5: 665-847.
- Reinthal, P. N. & S. M. Lewis. 1986. Social behaviour, foraging efficiency and habitat utilization in a group of tropical herbivorous fish. Animal Behaviour, 34: 1687-1693.
- Ribeiro, F. P. 2004. Composição da biocenose e abundância relativa de peixes capturados com covos nos estados do Rio Grande do Norte e Pernambuco (Brasil). Boletim Técnico-Científico do CEPENE, 12: 113-128.
- Risk, A. 1998. The effects of interactions with reef residents on the settlement and subsequent persistence of ocean surgeonfish, Acanthurus bahianus . Environmental Biology of Fishes, 51: 377-389.
- Roberts, G. 1996. Why individual vigilance declines as group size increases. Animal Behaviour, 51: 1077-1086.
- Rocha, L. A., A. L. Bass, D. R. Robertson & B. W. Bowen, 2002. Adult habitat preferences, larval dispersal, and the comparative phylogeography of three Atlantic surgeonfishes (Teleostei: Acanthuridae). Molecular Ecology, 11: 243-252.
- Rosenthal, S. B., C. R. Twomey, A. T. Hartnett, H. S. Wu & I. D. Couzin. 2015. Revealing the hidden networks of interaction in mobile animal groups allows prediction of complex behavioral contagion. Proceedings of the National Academy of Sciences, 112: 4690-4695.
- Sampaio, C. L. S. & M. C. Nottingham. 2008. Guia para identificação de peixes ornamentais brasileiros: espécies marinhas. Brasília, DF, IBAMA, v.1 (205p).
- Sampaio, C. L. S. & I. L. Rosa. 2005. A coleta de peixes ornamentais marinhos na Bahia, Brasil: técnicas utilizadas e implicações à conservação. Boletim Técnico-Científico do CEPENE, 13: 39-51.
- Sakai, Y. & M. Kohda. 1995. Foraging by mixed-species groups involving a small Angelfish, Centropyge ferrugatus (Pomacanthidae). Japonese Journal of Ichthyology, 41: 429-435.
- Sazima, I., R. L. Moura& J. L. Gasparini. 1998. The wrasse Halichoeres cyanocephalus (Labridae) as a specialized cleaner fish. Bulletin of Marine Science, 63: 605-610.
- Semeniuk, C. A. D. & L. M. Dill. 2005. Cost/benefit analysis of group and solitary resting in the cowtail stingray, Pastinachus sephen . Behavioral Ecology, 16: 417-426.
- Semeniuk, C. A. D. & L. M. Dill. 2006. Anti-predator benefits of mixed-species groups of cowtail stingrays (Pastinachus sephen ) and whiprays (Himantura uarnak ) at rest. Ethology, 112: 33-43.
- Smith, M. P. L., J. D. Bell, D. A. Pollard & B.C. Russel. 1989. Catch and effort of competition spearfishermen in Southeastern Australia. Fisheries Research, 8: 45-61.
- Stankowich, T. 2008. Ungulate flight responses to human disturbance: a review and meta-analysis. Biological Conservation, 141: 2159-2173.
- Stankowich, T. & D. T. Blumstein. 2005. Fear in animals: a meta-analysis and review of risk assessment. Proceedings of the Royal Society B: Biological Sciences, 272: 2627-2634.
- Stankowich, T. & R. G. Coss. 2007. Effects of risk assessment, predator behavior, and habitat on escape behavior in Columbian black-tailed deer. Behavioral Ecology, 18: 358-367.
- Witter, M. S., I. C. Cuthill & R. H. C. Bonser. 1994. Experimental investigations of mass-dependent predation risk in the European starling, Sturnus vulgaris . Animal Behaviour, 48: 201-222.
- Wolf, N. G. 1987. Schooling tendency and foraging benefit in the ocean surgeonfish. Behavioral Ecology and Sociobiology, 21: 59-63.
- Ydenberg, R. C. & L. M. Dill. 1986. The Economics of fleeing from predators. Advances in the Study of Behavior, 16: 229-249.
Publication Dates
-
Publication in this collection
2016
History
-
Received
25 May 2015 -
Accepted
04 Nov 2015