Abstract
The role of several feline vector-borne pathogens (FVBP) as a cause of disease in cats has not been clearly determined. In fact, with the exception of Bartonella spp. and hemoplasmas, FVBP in cats has not been clearly determined in Brazil yet. The present study aimed at identifying, by using molecular methods, the presence of FVBP in three cats showing non-specific clinical signs and inclusions suggestive of hemoparasites in blood smears. Cytauxzoon felis, ‘Candidatus Mycoplasma haemominutum’, Ehrlichia sp. closely related to Ehrlichia canis, and Anaplasma sp. closely related to Anaplasma phagocytophilum were detected in blood samples from two out of three sampled cats. Both cats positive for multiple FVBP did not show hematological and biochemical abnormalities. The present work emphasizes the need for molecular confirmation of co-infection by multiple FVBP in cats presenting non-specific clinical signs and inclusions resembling hemoparasites in blood smears.
Keywords:
Anaplasma; Cytauxzoon; Ehrlichia; Mycoplasma; feline
Resumo
O papel de diversos patógenos felinos transmitidos por vetores (PFTV) como causa de enfermidades em gatos não tem sido claramente determinado. De fato, com exceção de Bartonella spp. e hemoplasmas, PFTV têm sido bem menos estudados no Brasil. O presente estudo objetivou investigar, utilizando métodos moleculares, a presença de PFTV em três gatos apresentando sinais clínicos inespecíficos e inclusões sugestivas de hemoparasitas em esfregaços sanguíneos. Cytauxzoon felis, ‘Candidatus Mycoplasma haemominutum’, Ehrlichia sp. filogeneticamente relacionada a Ehrlichia canis, e Anaplasma sp. filogeneticamente relacionado a Anaplasma phagocytophilum foram detectados em amostras de sangue de dois dos três gatos amostrados. Os dois gatos positivos para múltiplos PFTV não apresentaram alterações hematológicas e bioquímicas. O presente trabalho enfatiza a necessidade de confirmação molecular da infecção por múltiplos PFTV em gatos apresentando sinais clínicos inespecíficos e inclusões sugestivas de hemoparasitas em esfregaços sanguíneos.
Palavras-chave:
Anaplasma; Cytauxzoon; Ehrlichia; Mycoplasma; felino
Vector-borne diseases have wide distribution and raising occurrence, mainly due to climatic and environmental changes and the increase of the mobility of people and animals. Such events may contribute to the multiplication and dispersion of vectors and pathogens, causing morbidity and mortality of animal and humans (BANETH et al., 2012Baneth G, Bourdeau P, Bourdoiseau G, Bowman D, Breitschwerdt E, Capelli G, et al. Vector-borne diseases–constant challenge for practicing veterinarians: recommendations from the CVBD World Forum. Parasit Vectors 2012; 5(1): 55. PMid:22433172. http://dx.doi.org/10.1186/1756-3305-5-55.
http://dx.doi.org/10.1186/1756-3305-5-55...
).
Cats, especially those wandering or living in public shelters, are at high risk of acquiring vector-borne pathogens (VBP), probably because they are often not treated with ectoparasiticides. In addition, the general conditions of these animals (eg., poor nutrition) may contribute to the susceptibility to VBP (OTRANTO & DANTAS-TORRES, 2010Otranto D, Dantas-Torres F. Canine and feline vector-borne diseases in Italy: current situation and perspectives. Parasit Vectors 2010; 3(1): 2. PMid:20145730. http://dx.doi.org/10.1186/1756-3305-3-2.
http://dx.doi.org/10.1186/1756-3305-3-2...
). Previous studies highlighted the circulation of several pathogens with zoonotic potential in cats in Brazil, emphasizing the need for a molecular approach in the identification of such agents (ANDRÉ et al., 2014André MR, Denardi NCB, Sousa KCM, Gonçalves LR, Henrique PC, Ontivero CRGR, et al. Arthropod-borne pathogens circulating in free-roaming domestic cats in a zoo environment in Brazil. Ticks Tick Borne Dis 2014; 5(5): 545-551. PMid:24889035. http://dx.doi.org/10.1016/j.ttbdis.2014.03.011.
http://dx.doi.org/10.1016/j.ttbdis.2014....
, 2015André MR, Herrera HM, Fernandes SJ, Sousa KCM, Gonçalves LR, Domingos IH, et al. Tick-borne agents in domesticated and stray cats from the city of Campo Grande, state of Mato Grosso do Sul, midwestern Brazil. Ticks Tick Borne Dis 2015; 6(6): 779-786. PMid:26187416. http://dx.doi.org/10.1016/j.ttbdis.2015.07.004.
http://dx.doi.org/10.1016/j.ttbdis.2015....
). Since most of the diseases caused by VBP show nonspecific clinical signs, which may vary according to the stage of infection, the molecular diagnosis favors the correct determination of the prognosis and the choice of an effective treatment (GONÇALVES et al., 2014Gonçalves LR, Filgueira KD, Ahid SMM, Pereira JS, Vale AM, Machado RZ, et al. Study on coinfecting vector-borne pathogens in dogs and ticks in Rio Grande do Norte, Brazil. Rev Bras Parasitol Vet 2014; 23(3): 407-412. PMid:25271465. http://dx.doi.org/10.1590/S1984-29612014071.
http://dx.doi.org/10.1590/S1984-29612014...
).
The present study aimed at identifying, by using molecular methods, the presence of FVBP (Anaplasma spp., Ehrlichia spp., Babesia spp., Bartonella spp., Mycoplasma spp., Cytauxzoon spp. and Hepatozoon spp.) in cats showing non-specific clinical signs and inclusions suggestive of hemoparasites in blood smears.
In March, 2016, EDTA-whole blood samples were collected from three cats showing non-specific clinical signs and attended at Veterinary Hospital of University Federal Rural do Semi Árido (UFERSA), Mossoró, Rio Grande do Norte, Brazil (5° 12’ 10” S, 37° 19’ 32” O).
A non-castrated male cat, 1 year and 2 months old (# Cat 1), arrived at the veterinary hospital with signs of pruritus, diarrhea and hyporexia. Clinical examination showed cephalic alopecia in the parietal, retroauricular and tail extremities, melena and thickening of the intestinal loops at abdominal palpation. The animal had access to the street with a history of fights with other cats. In the anamnesis, contact with birds, rabbits and dogs, was reported. The owner informed that while vaccination (triple viral against panleukopenia, rhinotracheitis and calicivirus, and anti-rabies) was updated, deworming was out of date. Although hematological and biochemical alterations were not observed, erythrocytes inclusions inside erythrocytes suggestive of piroplasmids were identified (Figure 1).

Piroplasmid-suggestive forms found in Panoptic-stained blood smears found within erythrocytes from a domestic cat from Mossoró, Rio Grande do Norte, Brazil.
In addition, a 3-year-old castrated female (#Cat2) presented anorexia, emesis, asthenia, accelerated weight loss, icteric mucosa, hypothermia, opaque fur, dehydration, submandibular lymphadenomegaly, and a firm bulging abdomen. It was an indoor cat living with two other cats. The owner informed that both vaccination and deworm were updated. Hematological examination revealed neutrophilia (15410/mm3), thrombocytopenia (90 mil/mm3), anisocytosis, hypochromia, moderate polychromasia, metarrubycytes, presence of Howell-Jolly corpuscles and presence of inclusions inside erythrocytes, suggestive of piroplasmids. In the biochemical analysis, while urea (19 mg/dL) and albumin (1.7 g/dL) were below the reference values, alkaline phosphatase (566 U/L), cholesterol (239 mg/dL) and triglycerides (1146 mg/dL) were above the reference values.
Finally, a 2 years and 2 months old, non-castrated male cat (# Cat 3), arrived at the veterinary hospital due to dehiscence of a surgical wound after an osteosynthesis of an exposed fracture in the right thoracic limb. The animal presented asthenia, claudication, edema, ulcer and purulent exudate with exposure of the musculature and bone tissue in the affected limb. The owner reported that the animal had no access to the street and no contact with other animals. Both vaccination (only anti-rabies) and deworm were updated. Despite the lack of hematological and biochemical alterations, inclusions inside erythrocytes suggestive of piroplasmids were found in Panotic-blood smears.
DNA was extracted from 200 µL of each cat whole blood using the QIAamp DNA Blood Mini kit (QIAGEN®, Valencia, CA, USA), according to the manufacturer’s instructions. DNA concentration and quality were measured using 260/280nm absorbance ratio (Nanodrop®, Thermo Fisher Scientific, Waltham, MA, USA). In order to verify the presence of amplifiable DNA in the samples, an internal control PCR assay targeting a fragment of mammalian glyceraldehyde-3-phosphatedehydrogenase (GAPDH) (BIRKENHEUER et al., 2003Birkenheuer AJ, Levy MG, Breitschwerdt EB. Development and evaluation of a seminested PCR for detection and differentiation of Babesia gibsoni (Asian genotype) and B. canis DNA in canine blood samples. J Clin Microbiol 2003; 41(9): 4172-4177. PMid:12958243. http://dx.doi.org/10.1128/JCM.41.9.4172-4177.2003.
http://dx.doi.org/10.1128/JCM.41.9.4172-...
) was performed. Conventional PCR (cPCR) assays targeting 16S rRNA gene of Ehrlichia spp. (MURPHY et al., 1998Murphy GL, Ewing SA, Whitworth LC, Fox JC, Kocan AA. A molecular and serologic survey of Ehrlichia canis, E. chaffeensis and E. ewingii in dogs and ticks from Oklahoma. Vet Parasitol 1998; 79(4): 325-339. PMid:9831955. http://dx.doi.org/10.1016/S0304-4017(98)00179-4.
http://dx.doi.org/10.1016/S0304-4017(98)...
), Anaplasma spp. (MASSUNG et al., 1998Massung RF, Slater K, Owens JH, Nicholson WL, Mather TN, Solberg VB, et al. Nested PCR assay for detection of granulocytic ehrlichiae. J Clin Microbiol 1998; 36(4): 1090-1095. PMid:9542943.), Mycoplasma spp. (16S rRNA) (MAGGI et al., 2013Maggi RG, Chitwood MC, Kennedy-Stoskopf S, DePerno CS. Novel hemotropic Mycoplasma species in white-tailed deer (Odocoileus virginianus). Comp Immunol Microbiol Infect Dis 2013; 36(6): 607-611. PMid:24018179. http://dx.doi.org/10.1016/j.cimid.2013.08.001.
http://dx.doi.org/10.1016/j.cimid.2013.0...
), and 18S rRNA of Cytauxzoon felis (18S rRNA) (BIRKENHEUER et al., 2006Birkenheuer AJ, Marr H, Alleman AR, Levy MG, Breitschwerdt EB. Development and evaluation of a PCR assay for the detection of Cytauxzoon felis DNA in feline blood samples. Vet Parasitol 2006; 137(1-2): 144-149. PMid:16417970. http://dx.doi.org/10.1016/j.vetpar.2005.12.007.
http://dx.doi.org/10.1016/j.vetpar.2005....
), Hepatozoon spp. (18S rRNA) (PERKINS & KELLER, 2001Perkins SL, Keller AK. Phylogeny of nuclear small subunit rRNA genes of hemogregarines amplified with specific primers. J Parasitol 2001; 87(4): 870-876. PMid:11534653. http://dx.doi.org/10.1645/0022-3395(2001)087[0870:PONSSR]2.0.CO;2.
http://dx.doi.org/10.1645/0022-3395(2001...
; UJVARI et al., 2004Ujvari B, Madsen T, Olsson M. High prevalence of Hepatozoon spp. (Apicomplexa, Hepatozoidae) infection in water pythons (Liasis fuscus) from tropical Australia. J Parasitol 2004; 90(3): 670-672. PMid:15270125. http://dx.doi.org/10.1645/GE-204R.
http://dx.doi.org/10.1645/GE-204R...
), Babesia spp. (18S rRNA) (JEFFERIES et al., 2007Jefferies R, Ryan UM, Irwin PJ. PCR-RFLP for the detection and differentiation of the canine piroplasm species and its use with filter paper-based technologies. Vet Parasitol 2007; 144(1-2): 20-27. PMid:17127005. http://dx.doi.org/10.1016/j.vetpar.2006.09.022.
http://dx.doi.org/10.1016/j.vetpar.2006....
) were performed. Additionally, a qPCR targeting a nuoG gene fragment of Bartonella spp. was also performed as previously described (ANDRÉ et al., 2015André MR, Herrera HM, Fernandes SJ, Sousa KCM, Gonçalves LR, Domingos IH, et al. Tick-borne agents in domesticated and stray cats from the city of Campo Grande, state of Mato Grosso do Sul, midwestern Brazil. Ticks Tick Borne Dis 2015; 6(6): 779-786. PMid:26187416. http://dx.doi.org/10.1016/j.ttbdis.2015.07.004.
http://dx.doi.org/10.1016/j.ttbdis.2015....
). Ehrlichia chaffeensis and Anaplasma phagocytophilum DNA samples, kindly supplied by Prof. John Stephen Dumler (Uniformed Services University of the Health Sciences, Bethesda, Maryland, USA), were used as positive controls in cPCR assays. The Jaboticabal strains of Ehrlichia canis and Babesia vogeli were obtained from experimentally infected dogs (CASTRO et al., 2004Castro MB, Machado RZ, Aquino LP, Alessi AC, Costa MT. Experimental acute canine monocytic ehrlichiosis: clinicopathological and immunopathological findings. Vet Parasitol 2004; 119(1): 73-86. PMid:15036578. http://dx.doi.org/10.1016/j.vetpar.2003.10.012.
http://dx.doi.org/10.1016/j.vetpar.2003....
; FURUTA et al., 2009Furuta PI, Oliveira TMFS, Teixeira MCA, Rocha AG, Machado RZ, Tinucci-Costa M. Comparison between a soluble antigen-based ELISA and IFAT in detecting antibodies against Babesia canis in dogs. Rev Bras Parasitol Vet 2009; 18(3): 41-45. PMid:19772774. http://dx.doi.org/10.4322/rbpv.01803007.
http://dx.doi.org/10.4322/rbpv.01803007...
). Finally, Hepatozoon spp., Cytauxzoon spp., Bartonella spp. and Mycoplasma spp. DNA positive controls were obtained from naturally infected animals (ANDRÉ et al., 2009André MR, Adania CH, Machado RZ, Allegretti SM, Felippe PAN, Silva KF, et al. Molecular detection of Cytauxzoon spp. in asymptomatic Brazilian wild captive felids. J Wildl Dis 2009; 45(1): 234-237. PMid:19204356. http://dx.doi.org/10.7589/0090-3558-45.1.234.
http://dx.doi.org/10.7589/0090-3558-45.1...
, 2010André MR, Adania CH, Teixeira RHF, Vargas GH, Falcade M, Sousa L, et al. Molecular detection of Hepatozoon spp. in Brazilian and exotic wild carnivores. Vet Parasitol 2010; 173(1-2): 134-138. PMid:20630658. http://dx.doi.org/10.1016/j.vetpar.2010.06.014.
http://dx.doi.org/10.1016/j.vetpar.2010....
, 2015André MR, Herrera HM, Fernandes SJ, Sousa KCM, Gonçalves LR, Domingos IH, et al. Tick-borne agents in domesticated and stray cats from the city of Campo Grande, state of Mato Grosso do Sul, midwestern Brazil. Ticks Tick Borne Dis 2015; 6(6): 779-786. PMid:26187416. http://dx.doi.org/10.1016/j.ttbdis.2015.07.004.
http://dx.doi.org/10.1016/j.ttbdis.2015....
). Ultra-pure water (Nuclease-FreeWater Promega®, Wisconsin, EUA) was used as negative control in all PCR assays. The gels were imaged under ultraviolet light using the Image Lab Software version 4.1 (Bio-Rad®).
The reaction products of positive samples in the PCR protocols above mentioned were purified using the Silica Bead DNA gel extraction kit (Thermo Fisher Scientific®, Waltham, MA, USA). Sanger sequencing was performed of both strands using the BigDye® Terminator v3.1Cycle Sequencing Kit (Thermo Fisher Scientific®, Waltham, MA, USA) and ABI PRISM 310DNA Analyzer (Applied Biosystems®, Foster City, CA, EUA) (SANGER et al., 1977Sanger F, Nicklen S, Coulson AR. DNA sequencing with chain-terminating inhibitors. Proc Natl Acad Sci USA 1977; 74(12): 5463-5467. PMid:271968. http://dx.doi.org/10.1073/pnas.74.12.5463.
http://dx.doi.org/10.1073/pnas.74.12.546...
). The sequences were aligned with sequences published in GenBank using MAFFT software, version 7) (KATOH & STANDLEY, 2013Katoh K, Standley DM. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol 2013; 30(4): 772-780. PMid:23329690. http://dx.doi.org/10.1093/molbev/mst010.
http://dx.doi.org/10.1093/molbev/mst010...
). Phylogenetic inference was based on Bayesian Inference (BI). The Bayesian inference (BI) analysis was performed with MrBayes 3.1.2 (RONQUIST & HUELSENBECK, 2003Ronquist F, Huelsenbeck JP. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003; 19(12): 1572-1574. PMid:12912839. http://dx.doi.org/10.1093/bioinformatics/btg180.
http://dx.doi.org/10.1093/bioinformatics...
). Markov chain Monte Carlo (MCMC) simulations were run for 109 generations with a sampling frequency of every 100 generations and a burn-in of 25%. The best model of evolution was selected by the program jModelTest2 (version 2.1.6) on XSEDE (DARRIBA et al., 2012Darriba D, Taboada GL, Doallo R, Posada D. jModelTest 2: more models, new heuristics and parallel computing. Nat Methods 2012; 9(8): 772. PMid:22847109. http://dx.doi.org/10.1038/nmeth.2109.
http://dx.doi.org/10.1038/nmeth.2109...
), under the Akaike Information Criterion (AIC) (POSADA & BUCKLEY, 2004Posada D, Buckley TR. Model selection and model averaging in phylogenetics: advantages of akaike information criterion and bayesian approaches over likelihood ratio tests. Syst Biol 2004; 53(5): 793-808. PMid:15545256. http://dx.doi.org/10.1080/10635150490522304.
http://dx.doi.org/10.1080/10635150490522...
). All phylogenetic analyses were performed using CIPRES Science Gateway (MILLER et al., 2011Miller MA, Pfeiffer W, Schwartz T. The CIPRES science gateway: a community resource for phylogenetic analyses [online]. New Orleans: CIPRES; 2011 [cited 2017 Jan 17]. Available from: https://www.phylo.org
https://www.phylo.org...
). The trees were examined in Treegraph 2.0.56-381 beta (STOVER & MULLER, 2010Stöver BC, Muller KF. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinformatics 2010; 11(1): 7. PMid:20051126. http://dx.doi.org/10.1186/1471-2105-11-7.
http://dx.doi.org/10.1186/1471-2105-11-7...
).
Cytauxzoon spp., Mycoplasma spp., Ehrlichia spp. and Anaplasma spp. DNA was detected in blood samples from two cats that presented non-specific clinical signs and structures suggestive of piroplasmids in Panoptic-stained blood smears (Table 1). The animal (#Cat 2) showing hematological and biochemical alterations were negative for all studied VBP. Babesia spp., Bartonella spp., and Hepatozoon spp. DNA were not detected in sampled cats’ blood samples.
Vector-borne pathogens molecularly detected in blood samples from three cats. Mossoró, Rio Grande do Norte, northeastern Brazil, 2016.
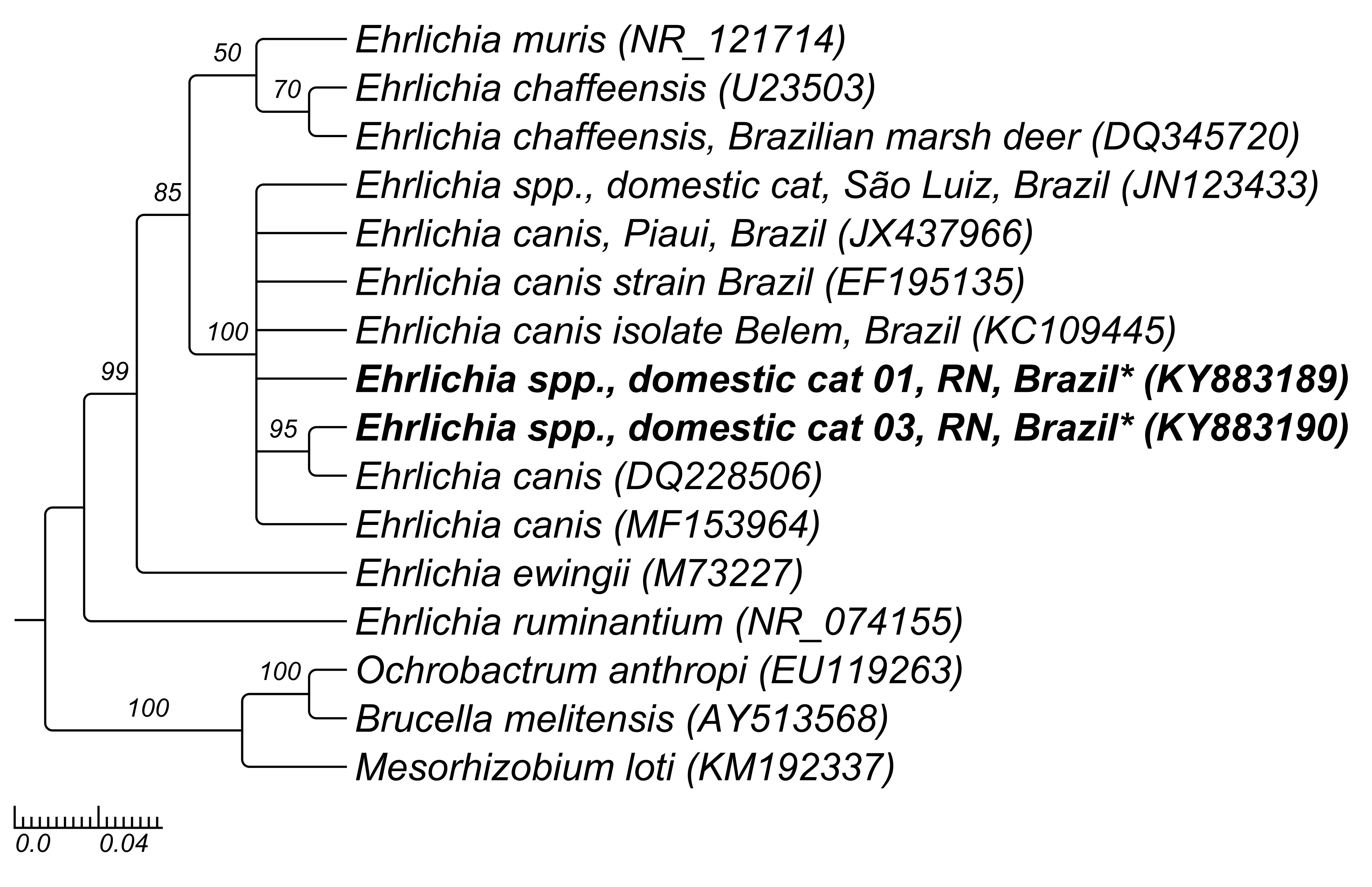
Concerning to phylogenetic analysis, the Anaplasma 16S rRNA sequences (MF288941-MF288942) detected in two cats were grouped with distinct Anaplasma sequences identified in a cat (KF964051) and in a Falconidae specimen (Caracara plancus - southern crested caracara) (JN217096) from Brazil; additionally, the sequence showed to be closely related to an A. phagocytophilum sequence (GU236670) detected in a dog from Austria, supported by a high posteriori probability (100%) in Bayesian inference (Figure 2). The Cytauxzoon sequence amplified in the present study was positioned nearest to C. felis sequence (L19080) supported by 88% index (Figure 3). The Ehrlichia 16S rRNA sequences amplified in the present study (KY883189 and KY883190) were positioned near to E. canis (JX437966; EF195135; KC109445) identified in dogs sampled from different Brazilian regions, as well as Ehrlichia sp. (JN123433) detected in a cat from São Luís, Maranhão state, and supported by a high posteriori probability (100%) in Bayesian analysis (Figure 4). Lastly, the Mycoplasma sequence amplified in our study was positioned closely to ‘Candidatus Mycoplasma haemominutum’ and supported by a high posteriori probability (100%) in Bayesian analysis (Figure 5).

Phylogenetic tree constructed with 600 pb Anaplasma spp.-16SrRNA sequences, using Bayesian method and GTR+G+I evolutionary model. Numbers at nodes correspond to Bayesian posterior probabilities over 50, using Mesorhizobium loti (KM192337), Brucella melitensis (AY513568) and Ochrobactrum anthropi (EU119263) as outgroups.

Phylogenetic tree constructed with 320 pb of Cytauxzoon-18SrRNA sequences, using Bayesian method and GTR+G+I evolutionary model. Numbers at nodes correspond to Bayesian posterior probabilities over 50, using Eimeria seviletensis (AF311644), Sarcocystis sp. (U97524), and Adelina bambarooniae (AF494059) as outgroups.

Phylogenetic tree constructed with 860 pb Ehrlichia spp.-16SrRNA sequences, using Bayesian method and GTR+G+I evolutionary model. Numbers at nodes correspond to Bayesian posterior probabilities over 50, using Mesorhizobium loti (KM192337), Brucella melitensis (AY513568) and Ochrobactrum anthropi (EU119263) as outgroups.

Phylogenetic tree constructed with 800pb Mycoplasma spp.16SrRNA sequences, using Bayesian method and GTR+G+I evolutionary model. Numbers at nodes correspond to Bayesian posterior probabilities over 50, using Mesorhizobium loti (KM192337), Brucella melitensis (AY513568) and Ochrobactrum anthropi (EU119263) as outgroups.
The role of several FVBP as a cause of disease in cats has not been clearly determined. In fact, with the exception of Bartonella spp. and hemotrophic mycoplasmas (ANDRÉ et al., 2014André MR, Denardi NCB, Sousa KCM, Gonçalves LR, Henrique PC, Ontivero CRGR, et al. Arthropod-borne pathogens circulating in free-roaming domestic cats in a zoo environment in Brazil. Ticks Tick Borne Dis 2014; 5(5): 545-551. PMid:24889035. http://dx.doi.org/10.1016/j.ttbdis.2014.03.011.
http://dx.doi.org/10.1016/j.ttbdis.2014....
, 2015André MR, Herrera HM, Fernandes SJ, Sousa KCM, Gonçalves LR, Domingos IH, et al. Tick-borne agents in domesticated and stray cats from the city of Campo Grande, state of Mato Grosso do Sul, midwestern Brazil. Ticks Tick Borne Dis 2015; 6(6): 779-786. PMid:26187416. http://dx.doi.org/10.1016/j.ttbdis.2015.07.004.
http://dx.doi.org/10.1016/j.ttbdis.2015....
), FVBP have been less frequently studied in Brazil and are generally under-estimated as a clinical entity in cats, as compared to dogs or people.
Feline infection with pathogens from order Rickettsiales is mostly associated with non-specific clinical signs, such as anorexia, lethargy, dehydration and fever (PENNISI et al., 2017Pennisi MG, Hofmann-Lehmann R, Radford AD, Tasker S, Belák S, Addie DD, et al. Anaplasma, Ehrlichia and Rickettsia species infections in cats: European guidelines from the ABCD on prevention and management. J Feline Med Surg 2017; 19(5): 542-548. PMid:28438088. http://dx.doi.org/10.1177/1098612X17706462.
http://dx.doi.org/10.1177/1098612X177064...
). In Brazil, the occurrence of Ehrlichia sp. closely related to E. canis has been already reported in cats sampled in the states of Minas Gerais (OLIVEIRA et al., 2009Oliveira LS, Mourão LC, Oliveira KA, Agostini MM, Oliveira AC, Almeida MR, et al. Molecular detection of Ehrlichia canis in cats in Brazil. Clin Microbiol Infect 2009;15(Suppl 2): 53-54. PMid:19374643. http://dx.doi.org/10.1111/j.1469-0691.2008.02175.x.
http://dx.doi.org/10.1111/j.1469-0691.20...
), Maranhão (BRAGA et al., 2012Braga MS, André MR, Freschi CR, Teixeira MC, Machado RZ. Molecular and serological detection of Ehrlichia spp. in cats on São Luís Island, Maranhão, Brazil. Rev Bras Parasitol Vet 2012; 21(1): 37-41. PMid:22534943. http://dx.doi.org/10.1590/S1984-29612012000100008.
http://dx.doi.org/10.1590/S1984-29612012...
), Mato Grosso (BRAGA et al., 2013Braga IA, Santos LG, Melo AL, Jaune FW, Ziliani TF, Girardi AF, et al. Hematological values associated to the serological and molecular diagnostic in cats suspected of Ehrlichia canis infection. Rev Bras Parasitol Vet 2013; 22(4): 470-474. PMid:24473870. http://dx.doi.org/10.1590/S1984-29612013000400005.
http://dx.doi.org/10.1590/S1984-29612013...
, 2014Braga IA, Santos LGF, Ramos DGS, Melo ALT, Mestre GLC, Aguiar DM. Detection of Ehrlichia canis in domestic cats in the central-western region of Brazil. Braz J Microbiol 2014; 45(2): 641-645. PMid:25242952. http://dx.doi.org/10.1590/S1517-83822014000200036.
http://dx.doi.org/10.1590/S1517-83822014...
) and Mato Grosso do Sul (ANDRÉ et al., 2015André MR, Herrera HM, Fernandes SJ, Sousa KCM, Gonçalves LR, Domingos IH, et al. Tick-borne agents in domesticated and stray cats from the city of Campo Grande, state of Mato Grosso do Sul, midwestern Brazil. Ticks Tick Borne Dis 2015; 6(6): 779-786. PMid:26187416. http://dx.doi.org/10.1016/j.ttbdis.2015.07.004.
http://dx.doi.org/10.1016/j.ttbdis.2015....
), and in wild felids maintained in captive in zoos in the state of São Paulo and Brasília (ANDRÉ et al., 2010André MR, Adania CH, Teixeira RHF, Vargas GH, Falcade M, Sousa L, et al. Molecular detection of Hepatozoon spp. in Brazilian and exotic wild carnivores. Vet Parasitol 2010; 173(1-2): 134-138. PMid:20630658. http://dx.doi.org/10.1016/j.vetpar.2010.06.014.
http://dx.doi.org/10.1016/j.vetpar.2010....
; 2012André MR, Dumler JS, Scorpio DG, Teixeira RHF, Allegretti SM, Machado RZ. Molecular detection of tick-borne bacterial agents in Brazilian and exotic captive carnivores. Ticks Tick Borne Dis 2012; 3(4): 247-253. PMid:22749737. http://dx.doi.org/10.1016/j.ttbdis.2012.04.002.
http://dx.doi.org/10.1016/j.ttbdis.2012....
). Although both cats positive in PCR assays for Ehrlichia sp. did not show any hematological alteration, low erythrocyte count, thrombocytopenia, lymphopenia, and monocytosis were previously reported in Ehrlichia-PCR positive cats in midwestern Brazil (BRAGA et al., 2013Braga IA, Santos LG, Melo AL, Jaune FW, Ziliani TF, Girardi AF, et al. Hematological values associated to the serological and molecular diagnostic in cats suspected of Ehrlichia canis infection. Rev Bras Parasitol Vet 2013; 22(4): 470-474. PMid:24473870. http://dx.doi.org/10.1590/S1984-29612013000400005.
http://dx.doi.org/10.1590/S1984-29612013...
).
Even though the positive cats sampled in the present study for Anaplasma sp. did not show any hematological disorders, neutrophilia with left shift, lymphopenia and thrombocytopenia have been found in A. phagocytophilum-naturally infected cats (BJÖERSDORFF et al., 1999Bjöersdorff A, Svendenius L, Owens JH, Massung RF. Feline granulocytic ehrlichiosis-a report of a new clinical entity and characterisation of the infectious agent. J Small Anim Pract 1999; 40(1): 20-24. PMid:10092038. http://dx.doi.org/10.1111/j.1748-5827.1999.tb03249.x.
http://dx.doi.org/10.1111/j.1748-5827.19...
; LAPPIN et al., 2004Lappin MR, Breitschwerdt EB, Jensen WA, Dunnigan B, Rha JY, Williams CR, et al. Molecular and serologic evidence of Anaplasma phagocytophilum infection in cats in North America. J Am Vet Med Assoc 2004; 225(6): 893-896. PMid:15485049. http://dx.doi.org/10.2460/javma.2004.225.893.
http://dx.doi.org/10.2460/javma.2004.225...
). In Brazil, genotypes closely related to A. phagocytophilum has been detected in stray cats São Paulo (ANDRÉ et al., 2014André MR, Denardi NCB, Sousa KCM, Gonçalves LR, Henrique PC, Ontivero CRGR, et al. Arthropod-borne pathogens circulating in free-roaming domestic cats in a zoo environment in Brazil. Ticks Tick Borne Dis 2014; 5(5): 545-551. PMid:24889035. http://dx.doi.org/10.1016/j.ttbdis.2014.03.011.
http://dx.doi.org/10.1016/j.ttbdis.2014....
) and in wild felids maintained in captivity (ANDRÉ et al., 2012André MR, Dumler JS, Scorpio DG, Teixeira RHF, Allegretti SM, Machado RZ. Molecular detection of tick-borne bacterial agents in Brazilian and exotic captive carnivores. Ticks Tick Borne Dis 2012; 3(4): 247-253. PMid:22749737. http://dx.doi.org/10.1016/j.ttbdis.2012.04.002.
http://dx.doi.org/10.1016/j.ttbdis.2012....
). Once infected, cats seem to remain chronic carriers for rickettsial agents (PENNISI et al., 2017Pennisi MG, Hofmann-Lehmann R, Radford AD, Tasker S, Belák S, Addie DD, et al. Anaplasma, Ehrlichia and Rickettsia species infections in cats: European guidelines from the ABCD on prevention and management. J Feline Med Surg 2017; 19(5): 542-548. PMid:28438088. http://dx.doi.org/10.1177/1098612X17706462.
http://dx.doi.org/10.1177/1098612X177064...
).
In the present study, two cats were infected by ‘Candidatus M. haemominutum’, the most frequent hemoplasma species found in cats, albeit showing a low pathogenicity (TASKER, 2010Tasker S. Haemotropic mycoplasmas: what’s their real significance in cats? J Feline Med Surg 2010; 12(5): 369-381. PMid:20417898. http://dx.doi.org/10.1016/j.jfms.2010.03.011.
http://dx.doi.org/10.1016/j.jfms.2010.03...
). This hemoplasma species frequently does not induce anemia, unless concurrent disease occurs (TASKER, 2010Tasker S. Haemotropic mycoplasmas: what’s their real significance in cats? J Feline Med Surg 2010; 12(5): 369-381. PMid:20417898. http://dx.doi.org/10.1016/j.jfms.2010.03.011.
http://dx.doi.org/10.1016/j.jfms.2010.03...
). In the present study, the two hemoplasma positive-cats were co-infected with other hemoparasites and the erythrocyte count was within the reference values.
Herein, two out of three cats presenting structures similar to piroplasms inside erythrocytes were positive for Cytauxzoon spp. The phylogenetic analysis positioned one obtained 18S rRNA sequence near to C. felis, corroborating previous studies involving wild felids (ANDRÉ et al., 2009André MR, Adania CH, Machado RZ, Allegretti SM, Felippe PAN, Silva KF, et al. Molecular detection of Cytauxzoon spp. in asymptomatic Brazilian wild captive felids. J Wildl Dis 2009; 45(1): 234-237. PMid:19204356. http://dx.doi.org/10.7589/0090-3558-45.1.234.
http://dx.doi.org/10.7589/0090-3558-45.1...
) and domesticated cats (MAIA et al., 2013Maia LMP, Cerqueira AMF, Macieira DB, Souza AM, Moreira NS, Silva AV, et al. Cytauxzoon felis and ‘Candidatus Mycoplasma haemominutum’ coinfection in a Brazilian domestic cat (Felis catus). Rev Bras Parasitol Vet 2013; 22(2): 289-291. PMid:23856727. http://dx.doi.org/10.1590/S1984-29612013000200049.
http://dx.doi.org/10.1590/S1984-29612013...
; ANDRÉ et al., 2015André MR, Herrera HM, Fernandes SJ, Sousa KCM, Gonçalves LR, Domingos IH, et al. Tick-borne agents in domesticated and stray cats from the city of Campo Grande, state of Mato Grosso do Sul, midwestern Brazil. Ticks Tick Borne Dis 2015; 6(6): 779-786. PMid:26187416. http://dx.doi.org/10.1016/j.ttbdis.2015.07.004.
http://dx.doi.org/10.1016/j.ttbdis.2015....
) in Brazil. The occurrence of Cytauxzoon sp. seems to be more frequent among wild felids than in domestic cats in Brazil (ANDRÉ et al., 2015André MR, Herrera HM, Fernandes SJ, Sousa KCM, Gonçalves LR, Domingos IH, et al. Tick-borne agents in domesticated and stray cats from the city of Campo Grande, state of Mato Grosso do Sul, midwestern Brazil. Ticks Tick Borne Dis 2015; 6(6): 779-786. PMid:26187416. http://dx.doi.org/10.1016/j.ttbdis.2015.07.004.
http://dx.doi.org/10.1016/j.ttbdis.2015....
). In fact, neotropical wild felids may act as reservoirs for species of family Theileriidae in South America (ANDRÉ et al., 2009André MR, Adania CH, Machado RZ, Allegretti SM, Felippe PAN, Silva KF, et al. Molecular detection of Cytauxzoon spp. in asymptomatic Brazilian wild captive felids. J Wildl Dis 2009; 45(1): 234-237. PMid:19204356. http://dx.doi.org/10.7589/0090-3558-45.1.234.
http://dx.doi.org/10.7589/0090-3558-45.1...
). Clinico-pathological findings associated to cytauxzoonosis, such as anemia, thrombocytopenia, leukopenia, hypoproteinemia, and hyperbilirrubinemia (SHERRILL & COHN, 2015Sherrill MK, Cohn LA. Cytauxzoonosis: diagnosis and treatment of an emerging disease. J Feline Med Surg 2015; 17(11): 940-948. PMid:26486980. http://dx.doi.org/10.1177/1098612X15610681.
http://dx.doi.org/10.1177/1098612X156106...
) were not found in the two Cytauxzoon-PCR positive cats. Therefore, we assumed that both cats might have survived to the schyzogonous phase, the most life-threatening phase of this parasite (WANG et al., 2017Wang JL, Li TT, Liu GH, Zhu XQ, Yao C. Two tales of Cytauxzoon felis infections in domestic cats. Clin Microbiol Rev 2017; 30(4): 861-885. PMid:28637681. http://dx.doi.org/10.1128/CMR.00010-17.
http://dx.doi.org/10.1128/CMR.00010-17...
). In this case, both cats may act as chronic carriers, playing a role as source of infection for arthropod vectors.
One animal (Cat#2) was negative by molecular testing in spite of showing suggestive intra-erythrocytic piroplasmids structures in blood smears. Although rapid diagnosis can be performed with thin blood smears, false positive results are often observed, since pleomorphic piroplasms may be mistaken as Howell-Jolly bodies, stain precipitate, water artifacts, or even hemoplasmas (WANG et al., 2017Wang JL, Li TT, Liu GH, Zhu XQ, Yao C. Two tales of Cytauxzoon felis infections in domestic cats. Clin Microbiol Rev 2017; 30(4): 861-885. PMid:28637681. http://dx.doi.org/10.1128/CMR.00010-17.
http://dx.doi.org/10.1128/CMR.00010-17...
), Babesia vogeli and Theileria spp. (ANDRÉ et al., 2014André MR, Denardi NCB, Sousa KCM, Gonçalves LR, Henrique PC, Ontivero CRGR, et al. Arthropod-borne pathogens circulating in free-roaming domestic cats in a zoo environment in Brazil. Ticks Tick Borne Dis 2014; 5(5): 545-551. PMid:24889035. http://dx.doi.org/10.1016/j.ttbdis.2014.03.011.
http://dx.doi.org/10.1016/j.ttbdis.2014....
, 2015André MR, Herrera HM, Fernandes SJ, Sousa KCM, Gonçalves LR, Domingos IH, et al. Tick-borne agents in domesticated and stray cats from the city of Campo Grande, state of Mato Grosso do Sul, midwestern Brazil. Ticks Tick Borne Dis 2015; 6(6): 779-786. PMid:26187416. http://dx.doi.org/10.1016/j.ttbdis.2015.07.004.
http://dx.doi.org/10.1016/j.ttbdis.2015....
). Therefore, the molecular confirmation of inclusions in feline erythrocytes is necessary in order to achieve a correct diagnosis and choose the correct therapy.
In the present study, co-infection by Ehrlichia, Anaplasma, Cytauxzoon and ‘Candidatus Mycoplasma haemominutum’ were detected in two cats. Previously, co-positivity for Ehrlichia/Anaplasma and Babesia was reported in cats in Portugal (MAIA et al., 2014Maia C, Ramos C, Coimbra M, Bastos F, Martins A, Pinto P, et al. Bacterial and protozoal agents of feline vector-borne diseases in domestic and stray cats from southern Portugal. Parasit Vectors 2014; 7(1): 115. PMid:24655431. http://dx.doi.org/10.1186/1756-3305-7-115.
http://dx.doi.org/10.1186/1756-3305-7-11...
). In Brazil, co-positivity for B. vogeli and ‘Candidatus Mycoplasma haemominutum’ was reported among stray cats in São Paulo (ANDRÉ et al., 2014André MR, Denardi NCB, Sousa KCM, Gonçalves LR, Henrique PC, Ontivero CRGR, et al. Arthropod-borne pathogens circulating in free-roaming domestic cats in a zoo environment in Brazil. Ticks Tick Borne Dis 2014; 5(5): 545-551. PMid:24889035. http://dx.doi.org/10.1016/j.ttbdis.2014.03.011.
http://dx.doi.org/10.1016/j.ttbdis.2014....
). A cat co-infected by and Cytauxzoon sp. and ‘Candidatus Mycoplasma haemominutum’ died in Rio de Janeiro (MAIA et al., 2013Maia LMP, Cerqueira AMF, Macieira DB, Souza AM, Moreira NS, Silva AV, et al. Cytauxzoon felis and ‘Candidatus Mycoplasma haemominutum’ coinfection in a Brazilian domestic cat (Felis catus). Rev Bras Parasitol Vet 2013; 22(2): 289-291. PMid:23856727. http://dx.doi.org/10.1590/S1984-29612013000200049.
http://dx.doi.org/10.1590/S1984-29612013...
). It is already known that co-infections with different canine vector-borne pathogens are quite frequent in dogs living in geographic areas where the presence of competent vectors overlaps (OTRANTO et al., 2009Otranto D, Dantas-Torres F, Breitschwerdt EB. Managing canine vector borne diseases of zoonotic concern: part one. Trends Parasitol 2009; 25(4): 157-163. PMid:19269898. http://dx.doi.org/10.1016/j.pt.2009.01.003.
http://dx.doi.org/10.1016/j.pt.2009.01.0...
). It seems like that this pattern is also valid for feline vector-borne pathogens. The effect of multiple arthropod-borne hemoparasites in the pathogenesis of feline vector-borne diseases warrants further investigation. Therefore, veterinarians should keep in mind that cats co-infected by several FVBP may present a non-characteristic, and even severe, clinical outcome, which will further complicate the diagnosis, treatment and prognosis (MAIA et al., 2014Maia C, Ramos C, Coimbra M, Bastos F, Martins A, Pinto P, et al. Bacterial and protozoal agents of feline vector-borne diseases in domestic and stray cats from southern Portugal. Parasit Vectors 2014; 7(1): 115. PMid:24655431. http://dx.doi.org/10.1186/1756-3305-7-115.
http://dx.doi.org/10.1186/1756-3305-7-11...
).
In conclusion, the present work emphasizes the need for molecular confirmation of co-infection by multiple vector-borne pathogens in cats presenting non-specific clinical signs and inclusions resembling hemoparasites in blood smears.
References
- André MR, Adania CH, Machado RZ, Allegretti SM, Felippe PAN, Silva KF, et al. Molecular detection of Cytauxzoon spp. in asymptomatic Brazilian wild captive felids. J Wildl Dis 2009; 45(1): 234-237. PMid:19204356. http://dx.doi.org/10.7589/0090-3558-45.1.234
» http://dx.doi.org/10.7589/0090-3558-45.1.234 - André MR, Adania CH, Teixeira RHF, Vargas GH, Falcade M, Sousa L, et al. Molecular detection of Hepatozoon spp. in Brazilian and exotic wild carnivores. Vet Parasitol 2010; 173(1-2): 134-138. PMid:20630658. http://dx.doi.org/10.1016/j.vetpar.2010.06.014
» http://dx.doi.org/10.1016/j.vetpar.2010.06.014 - André MR, Denardi NCB, Sousa KCM, Gonçalves LR, Henrique PC, Ontivero CRGR, et al. Arthropod-borne pathogens circulating in free-roaming domestic cats in a zoo environment in Brazil. Ticks Tick Borne Dis 2014; 5(5): 545-551. PMid:24889035. http://dx.doi.org/10.1016/j.ttbdis.2014.03.011
» http://dx.doi.org/10.1016/j.ttbdis.2014.03.011 - André MR, Dumler JS, Scorpio DG, Teixeira RHF, Allegretti SM, Machado RZ. Molecular detection of tick-borne bacterial agents in Brazilian and exotic captive carnivores. Ticks Tick Borne Dis 2012; 3(4): 247-253. PMid:22749737. http://dx.doi.org/10.1016/j.ttbdis.2012.04.002
» http://dx.doi.org/10.1016/j.ttbdis.2012.04.002 - André MR, Herrera HM, Fernandes SJ, Sousa KCM, Gonçalves LR, Domingos IH, et al. Tick-borne agents in domesticated and stray cats from the city of Campo Grande, state of Mato Grosso do Sul, midwestern Brazil. Ticks Tick Borne Dis 2015; 6(6): 779-786. PMid:26187416. http://dx.doi.org/10.1016/j.ttbdis.2015.07.004
» http://dx.doi.org/10.1016/j.ttbdis.2015.07.004 - Baneth G, Bourdeau P, Bourdoiseau G, Bowman D, Breitschwerdt E, Capelli G, et al. Vector-borne diseases–constant challenge for practicing veterinarians: recommendations from the CVBD World Forum. Parasit Vectors 2012; 5(1): 55. PMid:22433172. http://dx.doi.org/10.1186/1756-3305-5-55
» http://dx.doi.org/10.1186/1756-3305-5-55 - Birkenheuer AJ, Levy MG, Breitschwerdt EB. Development and evaluation of a seminested PCR for detection and differentiation of Babesia gibsoni (Asian genotype) and B. canis DNA in canine blood samples. J Clin Microbiol 2003; 41(9): 4172-4177. PMid:12958243. http://dx.doi.org/10.1128/JCM.41.9.4172-4177.2003
» http://dx.doi.org/10.1128/JCM.41.9.4172-4177.2003 - Birkenheuer AJ, Marr H, Alleman AR, Levy MG, Breitschwerdt EB. Development and evaluation of a PCR assay for the detection of Cytauxzoon felis DNA in feline blood samples. Vet Parasitol 2006; 137(1-2): 144-149. PMid:16417970. http://dx.doi.org/10.1016/j.vetpar.2005.12.007
» http://dx.doi.org/10.1016/j.vetpar.2005.12.007 - Bjöersdorff A, Svendenius L, Owens JH, Massung RF. Feline granulocytic ehrlichiosis-a report of a new clinical entity and characterisation of the infectious agent. J Small Anim Pract 1999; 40(1): 20-24. PMid:10092038. http://dx.doi.org/10.1111/j.1748-5827.1999.tb03249.x
» http://dx.doi.org/10.1111/j.1748-5827.1999.tb03249.x - Braga IA, Santos LG, Melo AL, Jaune FW, Ziliani TF, Girardi AF, et al. Hematological values associated to the serological and molecular diagnostic in cats suspected of Ehrlichia canis infection. Rev Bras Parasitol Vet 2013; 22(4): 470-474. PMid:24473870. http://dx.doi.org/10.1590/S1984-29612013000400005
» http://dx.doi.org/10.1590/S1984-29612013000400005 - Braga IA, Santos LGF, Ramos DGS, Melo ALT, Mestre GLC, Aguiar DM. Detection of Ehrlichia canis in domestic cats in the central-western region of Brazil. Braz J Microbiol 2014; 45(2): 641-645. PMid:25242952. http://dx.doi.org/10.1590/S1517-83822014000200036
» http://dx.doi.org/10.1590/S1517-83822014000200036 - Braga MS, André MR, Freschi CR, Teixeira MC, Machado RZ. Molecular and serological detection of Ehrlichia spp. in cats on São Luís Island, Maranhão, Brazil. Rev Bras Parasitol Vet 2012; 21(1): 37-41. PMid:22534943. http://dx.doi.org/10.1590/S1984-29612012000100008
» http://dx.doi.org/10.1590/S1984-29612012000100008 - Castro MB, Machado RZ, Aquino LP, Alessi AC, Costa MT. Experimental acute canine monocytic ehrlichiosis: clinicopathological and immunopathological findings. Vet Parasitol 2004; 119(1): 73-86. PMid:15036578. http://dx.doi.org/10.1016/j.vetpar.2003.10.012
» http://dx.doi.org/10.1016/j.vetpar.2003.10.012 - Darriba D, Taboada GL, Doallo R, Posada D. jModelTest 2: more models, new heuristics and parallel computing. Nat Methods 2012; 9(8): 772. PMid:22847109. http://dx.doi.org/10.1038/nmeth.2109
» http://dx.doi.org/10.1038/nmeth.2109 - Furuta PI, Oliveira TMFS, Teixeira MCA, Rocha AG, Machado RZ, Tinucci-Costa M. Comparison between a soluble antigen-based ELISA and IFAT in detecting antibodies against Babesia canis in dogs. Rev Bras Parasitol Vet 2009; 18(3): 41-45. PMid:19772774. http://dx.doi.org/10.4322/rbpv.01803007
» http://dx.doi.org/10.4322/rbpv.01803007 - Gonçalves LR, Filgueira KD, Ahid SMM, Pereira JS, Vale AM, Machado RZ, et al. Study on coinfecting vector-borne pathogens in dogs and ticks in Rio Grande do Norte, Brazil. Rev Bras Parasitol Vet 2014; 23(3): 407-412. PMid:25271465. http://dx.doi.org/10.1590/S1984-29612014071
» http://dx.doi.org/10.1590/S1984-29612014071 - Jefferies R, Ryan UM, Irwin PJ. PCR-RFLP for the detection and differentiation of the canine piroplasm species and its use with filter paper-based technologies. Vet Parasitol 2007; 144(1-2): 20-27. PMid:17127005. http://dx.doi.org/10.1016/j.vetpar.2006.09.022
» http://dx.doi.org/10.1016/j.vetpar.2006.09.022 - Katoh K, Standley DM. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol 2013; 30(4): 772-780. PMid:23329690. http://dx.doi.org/10.1093/molbev/mst010
» http://dx.doi.org/10.1093/molbev/mst010 - Lappin MR, Breitschwerdt EB, Jensen WA, Dunnigan B, Rha JY, Williams CR, et al. Molecular and serologic evidence of Anaplasma phagocytophilum infection in cats in North America. J Am Vet Med Assoc 2004; 225(6): 893-896. PMid:15485049. http://dx.doi.org/10.2460/javma.2004.225.893
» http://dx.doi.org/10.2460/javma.2004.225.893 - Maggi RG, Chitwood MC, Kennedy-Stoskopf S, DePerno CS. Novel hemotropic Mycoplasma species in white-tailed deer (Odocoileus virginianus). Comp Immunol Microbiol Infect Dis 2013; 36(6): 607-611. PMid:24018179. http://dx.doi.org/10.1016/j.cimid.2013.08.001
» http://dx.doi.org/10.1016/j.cimid.2013.08.001 - Maia C, Ramos C, Coimbra M, Bastos F, Martins A, Pinto P, et al. Bacterial and protozoal agents of feline vector-borne diseases in domestic and stray cats from southern Portugal. Parasit Vectors 2014; 7(1): 115. PMid:24655431. http://dx.doi.org/10.1186/1756-3305-7-115
» http://dx.doi.org/10.1186/1756-3305-7-115 - Maia LMP, Cerqueira AMF, Macieira DB, Souza AM, Moreira NS, Silva AV, et al. Cytauxzoon felis and ‘Candidatus Mycoplasma haemominutum’ coinfection in a Brazilian domestic cat (Felis catus). Rev Bras Parasitol Vet 2013; 22(2): 289-291. PMid:23856727. http://dx.doi.org/10.1590/S1984-29612013000200049
» http://dx.doi.org/10.1590/S1984-29612013000200049 - Massung RF, Slater K, Owens JH, Nicholson WL, Mather TN, Solberg VB, et al. Nested PCR assay for detection of granulocytic ehrlichiae. J Clin Microbiol 1998; 36(4): 1090-1095. PMid:9542943.
- Miller MA, Pfeiffer W, Schwartz T. The CIPRES science gateway: a community resource for phylogenetic analyses [online]. New Orleans: CIPRES; 2011 [cited 2017 Jan 17]. Available from: https://www.phylo.org
» https://www.phylo.org - Murphy GL, Ewing SA, Whitworth LC, Fox JC, Kocan AA. A molecular and serologic survey of Ehrlichia canis, E. chaffeensis and E. ewingii in dogs and ticks from Oklahoma. Vet Parasitol 1998; 79(4): 325-339. PMid:9831955. http://dx.doi.org/10.1016/S0304-4017(98)00179-4
» http://dx.doi.org/10.1016/S0304-4017(98)00179-4 - Oliveira LS, Mourão LC, Oliveira KA, Agostini MM, Oliveira AC, Almeida MR, et al. Molecular detection of Ehrlichia canis in cats in Brazil. Clin Microbiol Infect 2009;15(Suppl 2): 53-54. PMid:19374643. http://dx.doi.org/10.1111/j.1469-0691.2008.02175.x
» http://dx.doi.org/10.1111/j.1469-0691.2008.02175.x - Otranto D, Dantas-Torres F, Breitschwerdt EB. Managing canine vector borne diseases of zoonotic concern: part one. Trends Parasitol 2009; 25(4): 157-163. PMid:19269898. http://dx.doi.org/10.1016/j.pt.2009.01.003
» http://dx.doi.org/10.1016/j.pt.2009.01.003 - Otranto D, Dantas-Torres F. Canine and feline vector-borne diseases in Italy: current situation and perspectives. Parasit Vectors 2010; 3(1): 2. PMid:20145730. http://dx.doi.org/10.1186/1756-3305-3-2
» http://dx.doi.org/10.1186/1756-3305-3-2 - Pennisi MG, Hofmann-Lehmann R, Radford AD, Tasker S, Belák S, Addie DD, et al. Anaplasma, Ehrlichia and Rickettsia species infections in cats: European guidelines from the ABCD on prevention and management. J Feline Med Surg 2017; 19(5): 542-548. PMid:28438088. http://dx.doi.org/10.1177/1098612X17706462
» http://dx.doi.org/10.1177/1098612X17706462 - Perkins SL, Keller AK. Phylogeny of nuclear small subunit rRNA genes of hemogregarines amplified with specific primers. J Parasitol 2001; 87(4): 870-876. PMid:11534653. http://dx.doi.org/10.1645/0022-3395(2001)087[0870:PONSSR]2.0.CO;2
» http://dx.doi.org/10.1645/0022-3395(2001)087[0870:PONSSR]2.0.CO;2 - Posada D, Buckley TR. Model selection and model averaging in phylogenetics: advantages of akaike information criterion and bayesian approaches over likelihood ratio tests. Syst Biol 2004; 53(5): 793-808. PMid:15545256. http://dx.doi.org/10.1080/10635150490522304
» http://dx.doi.org/10.1080/10635150490522304 - Ronquist F, Huelsenbeck JP. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003; 19(12): 1572-1574. PMid:12912839. http://dx.doi.org/10.1093/bioinformatics/btg180
» http://dx.doi.org/10.1093/bioinformatics/btg180 - Sanger F, Nicklen S, Coulson AR. DNA sequencing with chain-terminating inhibitors. Proc Natl Acad Sci USA 1977; 74(12): 5463-5467. PMid:271968. http://dx.doi.org/10.1073/pnas.74.12.5463
» http://dx.doi.org/10.1073/pnas.74.12.5463 - Sherrill MK, Cohn LA. Cytauxzoonosis: diagnosis and treatment of an emerging disease. J Feline Med Surg 2015; 17(11): 940-948. PMid:26486980. http://dx.doi.org/10.1177/1098612X15610681
» http://dx.doi.org/10.1177/1098612X15610681 - Stöver BC, Muller KF. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinformatics 2010; 11(1): 7. PMid:20051126. http://dx.doi.org/10.1186/1471-2105-11-7
» http://dx.doi.org/10.1186/1471-2105-11-7 - Tasker S. Haemotropic mycoplasmas: what’s their real significance in cats? J Feline Med Surg 2010; 12(5): 369-381. PMid:20417898. http://dx.doi.org/10.1016/j.jfms.2010.03.011
» http://dx.doi.org/10.1016/j.jfms.2010.03.011 - Ujvari B, Madsen T, Olsson M. High prevalence of Hepatozoon spp. (Apicomplexa, Hepatozoidae) infection in water pythons (Liasis fuscus) from tropical Australia. J Parasitol 2004; 90(3): 670-672. PMid:15270125. http://dx.doi.org/10.1645/GE-204R
» http://dx.doi.org/10.1645/GE-204R - Wang JL, Li TT, Liu GH, Zhu XQ, Yao C. Two tales of Cytauxzoon felis infections in domestic cats. Clin Microbiol Rev 2017; 30(4): 861-885. PMid:28637681. http://dx.doi.org/10.1128/CMR.00010-17
» http://dx.doi.org/10.1128/CMR.00010-17
Publication Dates
-
Publication in this collection
09 Nov 2017 -
Date of issue
Oct-Dec 2017
History
-
Received
26 Sept 2017 -
Accepted
17 Oct 2017