Abstract
Most native Amazonian crops were domesticated in the periphery of the basin. The upper Madeira River basin is an important part of this periphery where several important crops were domesticated and others are suspected to have been domesticated or arrived early. Some of these crops have been reasonably well studied, such as manioc, peanut, peach palm, coca and tobacco, while others are not as well known, such as the hot peppers Capsicum baccatum and C. frutescens, and still others need confirmation, such as cocoyam and annatto. We review the information available for manioc, peach palm, Capsicum, peanut, annatto and cocoyam. The state-of-the-art for Capsicum frutescens, annatto and cocoyam is insufficient to conclude definitively that they were domesticated in the upper Madeira, while all the others have at least one of their origins or centers of diversity in the upper Madeira.
Keywords
Amazonian crops; Center of domestication; Crop domestication; Southwestern Amazonia
Resumo
A maioria dos cultivos nativos da Amazônia foi domesticada na periferia da bacia. A bacia do alto rio Madeira é uma parte importante dessa periferia, onde se suspeita que vários cultivos importantes foram domesticados, alguns são confirmados e outros possivelmente chegaram há muito tempo na região. Alguns destes cultivos foram razoavelmente bem estudados, tais como mandioca, amendoim, pupunha, coca e tabaco, enquanto outros não são tão bem conhecidos, como as pimentas Capsicum baccatum e C. frutescens, e ainda outros precisam de confirmação, como taioba e urucum. Revisamos as informações disponíveis para a mandioca, pupunha, Capsicum, amendoim, urucum e taioba. O estado da arte de Capsicum frutescens, urucum e taioba é insuficiente para concluir definitivamente que eles foram domesticados no alto rio Madeira, enquanto todos os outros têm pelo menos uma das suas origens ou centros de diversidade na região.
Palavras-chave
Cultivos amazônicos; Centro de domesticação; Domesticação de cultivos; Sudoeste da Amazônia
INTRODUCTION
The Madeira River drains a large portion of southwestern Amazonia, with a complex of headwater rivers that arise either in the Andes, hence with sediment-rich ‘white’ waters, or the western edge of the Brazilian shield, with sediment-poor ‘clear’ or ‘black’ waters. Soils vary from nutrient-rich volcanic soils in the foothills to less nutrient-rich Ultisols in the main basin, with nutrientrich floodplain soils along the white-water rivers. In the upper basin, vegetational types vary from closed canopy humid forests, to open canopy sub-humid forests, to scrub forests to savanna, with the Llanos de Mojos an especially important part of the region. This ecological complexity offered numerous useful plants to arriving hunter-gathers (Lombardo et al., 2013), some of which were domesticated in various parts of the upper basin over the next millennia.
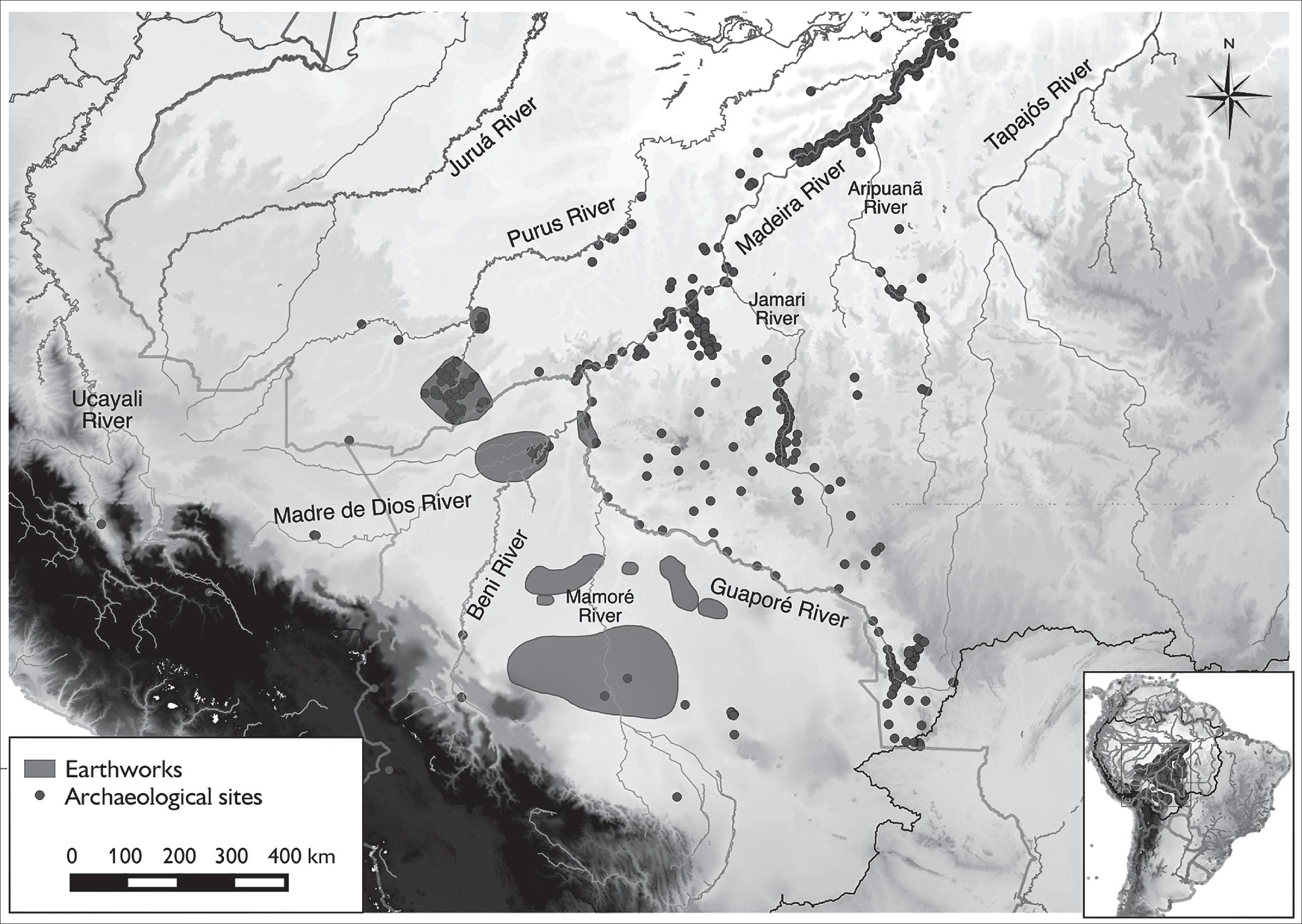
The arrival of humans in lowland South America may have occurred as early as 20,000 years before present (BP) (Lahaye et al., 2013), but surely by 13,000 BP (Roosevelt, 2013). Pre-ceramic societies occupied the area of Porto Velho, Rondônia, as early as 12,000 BP (Miller, 1992a), Lombardo et al. (2013) report initially preceramic shell mounds in the Llanos de Mojos, lowland Bolivia, dated to 10,600 BP, and Neves et al. (2015) are excavating a similar mound along the Guaporé River, Rondônia, so we can accept early occupation of the upper Madeira River basin as confirmed. This is important because some of the species that originated there may have started to be domesticated in this time frame. The first still pre-ceramic societies that were sedentary enough to generate terra preta de índio, or Amazonian Dark Earths, appeared by 4,800 BP along the Jamari River, not far from Porto Velho (Miller, 1992b), and along the Guaporé River (Neves et al., 2015) (Figure 1). Over the next millennia the upper Madeira River basin became the home of numerous complex societies who were responsible for various kinds of earthworks (raised fields, habitation mounds, geoglyphs, dike and canals etc.) (Dickau et al., 2012) and spoke numerous languages (Eriksen, 2011). The upper Madeira is the homeland of Tupi (Eriksen and Galucio, 2014) and is adjacent to the probable origin of Arawak in the upper Purus River (Walker and Ribeiro, 2011; Eriksen and Danielsen, 2014), the two most expansive language families of South America. The archaeobotany of the region is just starting to be explored with modern methods (Dickau et al., 2012; Whitney et al., 2014; Watling et al., 2015), although the species we will examine are seldom reported to date. Better preservation occurs along the dry Pacific coast of Peru, where many of these species are reported quite early (Pearsall, 1992).
Map of the upper Madeira River basin in southwestern Amazonia, highlighting known earthwork areas and other archaeological sites (extracted from Clement et al., 2015, with assistance from André Braga Junqueira, Wageningen University).
The periphery of Amazonia appears to be where the majority of Amazonian crops were domesticated (Clement et al., 2010). The upper Madeira River basin, hereafter upper Madeira, is an important part of the periphery and has been recognized as a probable or confirmed region of crop origins for some time, e.g., Piperno and Pearsall (1998; see also Piperno, 2011), who mentioned two hot peppers (Capsicum baccatum, C. pubescens), three legume seeds (Arachis hypogaea, Canavalia plagiosperma, Phaseolus vulgaris), and cocoyam (Xanthosoma sagittifolium). The Andean foothills and immediately adjacent lowlands of the upper Madeira also provided important drugs, e.g., coca (Erythroxylum coca; Plowman, 1984) and tobacco (Nicotiana tabacum; Gerstel and Sisson, 1995). The major root crop of Amazonia, manioc (Manihot esculenta) was confirmed to have originated in the upper Madeira (Schaal et al., 2006) and new information suggests the same for peach palm (Bactris gasipaes) (Cristo-Araújo et al., 2013). Clement et al. (2010) had modified Schultes’ hypothesis (1984) about annatto (Bixa orellana) having originated in Acre to include the upper Madeira, since wild annatto is found frequently in Rondônia (Moreira et al., 2015); however, these authors did not confirm the upper Madeira as the origin.
Over the millennia between crop origins and European conquest, crops were dispersed from the upper Madeira as far as Mexico, e.g., manioc, peanut, tobacco and annatto (Brown, 2010), and others arrived from there, e.g., maize (Zea mays) (Piperno, 2011), and elsewhere. At the time of European conquest, the complex societies of the Llanos de Mojos and adjacent areas of the upper Madeira cultivated numerous crops and certainly managed hundreds of species, leading Clement (1999) to propose a micro-center of diversity of crop genetic resources in the Llanos de Mojos (Clement et al., 2010). A micro-center is a geographically restricted area with an abundance of crop genetic resources used in both agroecosystems and in other domesticated landscapes. In this contribution we will review principally the genetic evidence, but also some morphology, biogeography and ethnography, for manioc, peach palm, Capsicum peppers, especially C. baccatum and C. frutescens, cocoyam and annatto, some of which originated in the upper Madeira and others of which were produced there before European conquest.
MANIOC
Manioc (Manihot esculenta Crantz, Euphorbiaceae) is grown throughout the tropics, and it is the primary source of carbohydrates for about 800 million people (Lebot, 2009). The origin of manioc as a crop has long been debated; it was once considered a cultigen (without a wild conspecific ancestral population) and was thought to have originated from a series of introgression events among wild species (Rogers and Appan, 1973). Based on phenotypic traits, Allem (2001) proposed that manioc consists of three subspecies: Manihot esculenta ssp. esculenta (cultivated form), M. esculenta ssp. flabellifolia (the closest wild relative) and M. esculenta ssp. peruviana (probably not involved in manioc domestication). Together with M. pruinosa, these form the primary gene pool of manioc.
The relationships among cultivated varieties of manioc and wild populations of M. esculenta ssp. flabellifolia and M. pruinosa were investigated with three different molecular markers [sequencing of the nuclear gene glyceraldehyde 3-phosphate dehydrogenese (G3pdh; Olsen and Schaal, 1999), Simple Sequence Repeats (Olsen and Schaal, 2001), and Single Nucleotide Polymorphisms (Olsen, 2004)]. In all cases, the genetic variability found in cultivated manioc is a subset of the genetic variability found in the populations of ssp. flabellifolia that occur in and adjacent to the upper Madeira (Schaal et al., 2006). These genetic results strongly suggest that manioc was domesticated only once from ssp. flabellifolia populations occurring in the upper Madeira of Mato Grosso and Rondônia states, and adjacent areas in the upper Purus and upper Juruá Rivers, Acre state, Brazil, and certainly in adjacent areas of Bolivia that were not sampled.
Divergent selective pressures gave origin to the two major groups of currently cultivated varieties: “sweet” manioc and “bitter” manioc (Mühlen et al., 2000; Elias et al., 2004). Sweet varieties have low amounts of cyanogenic glycosides (< 100 ppm fresh weight) and can be safely consumed with simple processing (cooking or sometimes even raw). On the other hand, bitter varieties have large amounts of cyanogenic glycosides (> 100 ppm fresh weight) and demand considerable detoxification and processing before consumption. Although this classification is dichotomous, continuous variation among manioc varieties and related wild species is observed, suggesting that sweet and bitter manioc are derived from an ancestor with intermediate toxicity (McKey and Beckerman, 1993). However, populations of ssp. flabellifolia in the interfluve between the central Guaporé River and the upper Machado River, in Rondônia, centrally located in the center of origin of domestication, are toxic (Perrut-Lima et al., 2014).
New data on the patterns of the distribution of genetic variation of the largest sweet and bitter manioc Brazilian sampling casts new light on the diffusion of the crop after its initial domestication (Mühlen et al., 2013). Although this study lacks manioc samples from the center of origin in Rondônia, the two major groups of cultivated manioc showed somewhat different patterns of diffusion across Brazilian ecogeographic regions. Sweet manioc appears to have diffused first from the center of origin, while bitter manioc appears to have originated later in a more central area of Amazonia, as hypothesized by Arroyo-Kalin (2010). Curiously, the Native Amazonians who first considered manioc to be important enough to have a name that can be reconstructed today were the Southern Arawak, of the Peruvian Amazonian lowlands in the upper Ucayali River, whose name for manioc reconstructs to 4,400 BP (Brown et al., 2013a), while the Tupi, who are closer to the origin in the eastern upper Madeira, are later (3,500 BP). However, manioc is thought to have originated as a domesticate much earlier, probably by 10,000 BP, given early archaeological finds in northwestern South America by 8,500 BP (Isendahl, 2011), which eliminates the Arawak as the original domesticators.
PEACH PALM
The peach palm (Bactris gasipaes Kunth, Palmae) is the only Neotropical palm with domesticated populations (Clement, 1988). It may have been selected initially for its wood, preferred for tool making, and later for its abundant oily fruits, and finally for starchiness in the fruits, making them good for fermentation (Clement et al., 2009a). The species presents considerable morphological and genetic variability in its wild and cultivated populations, due to adaptation to different environments and different stages of domestication, respectively. Thousands of years of domestication have resulted in landraces, each of which has common morphological, chemical and productive characteristics due to a common genetic origin (Mora Urpí et al., 1997). These landraces are widely distributed in the humid Neotropics, especially Amazonia.
Throughout the 20th century, peach palm was considered a cultigen, with no wild ancestor. The revision of Bactris (Henderson, 2000) gathered all cultivated populations of peach palm into var. gasipaes and all wild populations (previously identified as species) into var. chichagui (H. Karsten) Henderson, permitting phylogenetic hypotheses that can be tested with genetic tools. Within var. chichagui Henderson proposed three types, examined in detail by Clement et al. (2009b), who proposed that type 3 is not truly wild, but is the incipient domesticate.
There is considerable speculation about the origin of the founder event(s) that lead to domesticated peach palm populations, with three hypotheses under consideration until recently: a) a single domestication event in southwestern Amazonia (Clement, 1995), with some morphological (Ferreira, 1999) and molecular (Random Amplified Polymorphic DNA, a dominant marker; Rodrigues et al., 2004a) evidence, and the sympatry of one wild type (type 1) and the incipient domesticate (type 3); b) a single domestication event in northwestern South America, with archaeological coincidences (Morcote-Rios and Bernal, 2001), but with the occurrence of the incipient domesticate (type 3); and c) multiple domestication events in the distribution of the wild type 1 and the incipient domesticate (type 3), with coincidences in common SSR allele frequencies between var. chichagui and var. gasipaes in some localities (Mora-Urpí, 1999; Hernández-Ugalde et al., 2008, 2011).
Several genetic studies have been conducted over the last decade to understand the great genetic variability within and among landraces, and are reviewed in Clement et al. (2010). During the last decade, numerous microsatellite primers were developed and tested (Martínez et al., 2002; Billotte et al., 2004; Rodrigues et al., 2004b). Four of the first set were used to examine relationships among seven wild and eleven cultivated populations (Hernández-Ugalde et al., 2008, 2011). Unfortunately, they did not analyze the cultivated and wild populations separately, and created a neighbor-joining dendrogram from Nei’s minimum genetic distance, which they interpreted as showing three domestication events, even though the topology of the dendrogram was not much different from the first RAPD study (Rodrigues et al., 2004a). Although quite intriguing, the small number of microsatellites does not permit much precision.
A study by Reis (2009) examined the relationships within the landrace complex represented in the newly created core collection (Cristo-Araújo et al., 2015), which has 40 accessions, with 17 microsatellite loci. This analysis used the program Structure (Pritchard et al., 2000) to revalidate landraces and Nei’s (1978) genetic distance to create a neighbor-joining dendrogram, which was quite similar to the first and subsequent RAPD dendrograms (Rodrigues et al., 2004a; Cristo-Araújo et al., 2010), as well as the first microsatellite dendrogram (Hernández-Ugalde, 2005). The analyses with nuclear microsatellites confirmed previous analyses that showed a deep split between the eastern Amazonian landraces, on the one hand, and western Amazonian to Central American landraces, on the other (Rodrigues et al., 2004a; Cristo-Araújo et al., 2010; Hernández-Ugalde et al., 2011). As mentioned, however, these markers alone are insufficient for determining if the split represents two or more domestication events, as suggested by Mora Urpí (1999) and Hernández-Ugalde et al. (2011), or one event with two dispersal routes, as suggested by Rodrigues et al. (2004a) and Cristo-Araújo et al. (2010).
A universal chloroplast DNA sequence (Shaw et al., 2007) was used to determine the phylogenetic relationships among cultivated and wild populations in the core collection, as well as the closely related B. riparia and the more distantly related B. simplicifrons, both used as outgroups. The chloroplast haplotypes distinguish the eastern and southwestern cultivated populations from the western populations by a 12 base pair deletion (Cristo-Araújo et al., 2013), supporting the deep divergence among these groups observed by all studies with nuclear markers. Most important, however, the wild var. chichagui type 1 from Rio Branco, Acre, adjacent to the upper Madeira, contains both the eastern and the western chloroplast haplotypes, and groups with cultivated populations from Plácido de Castro, Acre, and Madre de Dios, Peru, both in the upper Madeira (Cristo-Araújo et al., 2013).
These new analyses suggest a single domestication event in southwestern Amazonia, probably in the upper Madeira, although the upper Ucayali River cannot yet be ruled out. After domestication, cultivated peach palm was dispersed down the Ucayali River and throughout western Amazonia, northwestern South America and southern Central America. At the same time, but perhaps later, because the fruits are less derived, cultivated peach palm was dispersed along the Madeira River to eastern Amazonia.
CAPSICUM PEPPERS
The genus Capsicum (Solanaceae) is represented by an extremely variable group of sweet and, principally, hot peppers. By the time of European conquest the chili peppers were present from northern Chile and Argentina, throughout the Andean and lowland South American countries, through Central America to northern Mexico and the southern United States (Barbieri and Neitzke, 2008; Pickersgill, 1969). Today they are distributed worldwide, and grown in temperate and tropical areas for use as spices, vegetables, ornamental plants, medicine and even defense (Reifschneider, 2000), and represent one of the most economically important vegetable crops worldwide (Albrecht et al., 2012).
Different species of Capsicum were domesticated independently in several regions of the Americas (Heiser Jr., 1995; Pickersgill, 2007): Mesoamerica (C. annuum, possibly C. frutescens), the mid-elevation Andes (C. pubescens), the mid-to-low elevation Andes and immediately adjacent lowlands of Bolivia and Peru (C. baccatum) (Albrecht et al., 2012), and the tropical lowlands of South America (C. chinense and possibly C. frutescens) (Pickersgill, 2007). The first peoples who considered Capsicum peppers important enough to be clearly named were the southern Arawak (Brown et al., 2013b) in the Amazonian lowlands of southern Peru, suggesting that C. baccatum may have been involved since it was domesticated in the northwestern upper Madeira and adjacent southern upper Ucayali (Albrecht et al., 2012). In each area of origin, one species was brought into domestication and they were then dispersed to different areas where they continued to be selected, resulting in distinct morphological types. Domestication resulted in change, especially in the fruits. The fruits of the wild types are small, erect, red and deciduous, while fruits of domesticates are larger, often pendent, not deciduous and varied in color. Domestication also resulted in changes in reproduction and the level of pungency (Heiser Jr., 1995).
During our recent expeditions in Brazilian Amazonia four of these domesticated species were found (M. Cristo-Araújo, personal observation): 1) C. annuum (chili, cayenne) was found in Roraima and in Southern Amazonia from Humaitá to Rondônia, along the upper Madeira, but not along the Solimões River; 2) C. baccatum (girl’s finger, chili, ají) was found only along the Mamoré River in Rondônia, close to its origin in northwestern Bolivia (Albrecht et al., 2012); 3) C. frutescens (malagueta, tabasco) presented very little variation in fruit appearance (size, shape, color) and was found along the north-south (Roraima-Rondônia) axis, especially along the upper Madeira, and in lower abundance along the Solimões River; 4) C. chinense (murupi, cumari, biquinho) was abundant along the Solimões River, in Roraima, in the northern Madeira-Purus interfluve and, in greater abundance, in the upper Madeira. In all localities visited there were more plants of C. chinense, with great variability in size, pungency, color and shape of the fruits. Some plants presented characteristics of two species and appeared to be hybrids, most likely C. chinense x C. frutescens (B. Pickersgill, personal communication). An apparently wild sample of C. chinense was found in Roraima (Barbosa et al., 2002), offering the possibility of a more precise origin for this species. However, no wild populations of C. frutescens have yet been found in Amazonia (or elsewhere), although the cultivated form is abundant in the upper Madeira, which is suggestive, but not proof.
PEANUT
The peanut (Arachis hypogaea L., Fabaceae) was domesticated for its seed, one of the few producers of underground seeds among cultivated plants. The genus Arachis is composed of 80 species (Krapovickas and Gregory, 1994; Valls and Simpson, 2005), with A. hypogaea the best-known and most widely dispersed in the Neotropics before conquest (Fávero and Valls, 2009). Arachis hypogaea contains two subspecies and six varieties: subsp. hypogaea, var. hypogaea and var. hirsuta; subsp. fastigiata, var. aequatoriana, var. fastigiata, var. peruviana and var. vulgaris (Krapovickas and Gregory, 1994). A recent molecular genetic study used a chloroplast sequence and the non-transcribed spacer of the nuclear 5S rDNA to decipher the origin of cultivated peanut (Grabiele et al., 2012): A. duranensis and A. ipaensis hybridized and gave rise to the wild tetraploid A. monticola, in northern Argentina. A. hypogaea was selected from A. monticola in northern Argentina and southern Bolivia, from where it was dispersed and diversified into two subspecies with six varieties, providing a clear example of a domestication event followed by diversification, as outlined by Meyer and Purugganan (2013).
The domestication of the peanuts must have started 6000-7000 BP (Fávero and Valls, 2009). The oldest archaeological peanut record is from the Huarmey valley, near the Peruvian coast, dating to 3,500-4,500 years BP (Pearsall, 1992), although it is not clear which subspecies and variety is involved. Several diversification events occurred across the savanna regions of lowland South America and adjacent open forests south of Amazonia, as well as adjacent to the Atlantic Forest, leading to the centers of diversity identified by Krapovickas (1995). In the western Llanos de Mojos, subsp. fastigiata var. peruviana was especially diverse, while in the extreme south of the upper Madeira subsp. hypogaea var. hypogaea presented a diversity of runner, bunch and erect types (Fávero and Valls, 2009), suggesting the great importance of both subspecies in the upper Madeira.
ANNATTO
Annatto (Bixa orellana L., Bixaceae) is a shrubby plant domesticated in the Neotropics as a colorant used in many different cultural contexts for the red-orange pigment in the arils around its seeds (León, 2000). Domesticated populations of annatto were well distributed throughout the Neotropics at the time of conquest (Donkin, 1974). Its common name in Portuguese (urucum) comes from Tupi-guaraní languages (Balée, 2000) and means red, while the common name achiote in Central America comes from achiotl in Nahuatl (Ocampo, 1983). In Mexico it was one of the ingredients used in the preparation of pre-conquest chocolate, a drink “thickened with maize, heated with chilies, spiced with vanilla, tinted with achiote” (Norton, 2008).
For many years, B. excelsa, a large tree found in Southern Amazonia, was accepted as its wild ancestor (Ducke, 1946; Schultes, 1984; Clement et al., 2010). Recently, spontaneous populations of annatto, that should be called B. orellana var. urucurana (Kuntze, 1925), were identified as the wild ancestor (Moreira et al., 2015). The variety urucurana occurs in open forests or anthropogenic landscapes, although never cultivated, and always associated with riparian environments. While cultivated annatto always produces abundant pigment around its seeds, the variety urucurana contains variable amounts. In areas where they co-exist, gene flow between them results in changes of pigment production, especially in the cultivated types. The exact location where annatto was domesticated is still unclear because of the wide distribution of variety urucurana in northern South America.
In southwestern Amazonia, annatto use has long been present for dying cotton (Lévi-Strauss, 2004), but it also has symbolic values in passage rites and a painted body is associated with vitality, beauty and fertility among the Kaxinawa and Suruwaha people (Lagrou, 2006; Aparicio, 2015). Ethnographies describe annatto as a plant of superior quality because it arose from the ashes, which suggests selection practices and fire management during the process of annatto domestication (Lévi-Strauss, 2004). The Nambicuara, of the Guaporé River basin, cultivated annatto in homegardens along the bluffs after burning and opening gallery forests (Lévi-Strauss, 1948), while young individuals of annatto were also transplanted near to home (Lévi-Strauss, 1957). The only archaeological record of annatto in Amazonia comes from the Llanos de Mojos, in lowland Bolivia, dated to 2400 years before present (BP) (Erickson, 1995), but there are older dates in the Caribbean, suggesting Arawak dispersal (Moreira et al., 2015).
COCOYAMS
Cocoyams or taiobas (Xanthosoma spp, Araceae) are perennial herbs, some with edible corms, others with edible leaves and still others used for medicinal purposes (Bown, 2000; Lebot, 2009). The name tajá or taiá is popularly used in Brazil as a generic term for Araceae species (Sauer, 1987; Silva, 2007) and is synonymous with taioba in Amazonian legends and myths (Martins and Teixeira, 2006). Wild types have high levels of calcium oxalate crystals in the corms that irritate the mucous membranes of the mouth, hence the side corms used must be dried, cooked or fermented to be eaten (Bown, 2000). There is linguistic evidence of this crop in Central America since 3000 BP (Hawkes, 1989). In South America, taioba pollen grains were registered in raised field archaeological sites in the Llanos de Mojos in the upper Madeira (Erickson, 1995).
The taxonomy of the genus Xanthosoma is unclear (Plucknet, 1976; Lebot, 2009). There are some 40 types of cultivated cocoyams, but in general these are lumped together as X. sagittifolium (Bown, 2000). All of them have 26 chromosomes and differ by pigmentation of leaves, plant height, shape, color and number of corms. These morphological features vary depending on environmental conditions (Lebot, 2009), suggesting significant plasticity (genotype by environment interaction). Traditionally, cocoyams are cultivated on low mounds mulched with dead leaves and grass with sufficient water for growth (Bown, 2000), although they are more tolerant of insufficient water than taro (Colocasia esculenta). Molecular analysis based on RAPD of two cultivated inter-fertile species (X. violaceum and X. caracu) from the West Indies showed low genetic variation (Jacobson et al., 1999), which suggests that some recognized species of Xanthosoma may be just clones.
It is still impossible to determine a center of origin of domesticated populations, especially because wild congenerics occur in both Central and South America (Piperno, 2011). It is possible that clones selected in Central America could have been introduced quite early (e.g., 3400 BP) into South America (Colunga-GarcíaMarín and Zizumbo-Villarreal, 2004) and Purseglove (1972) argued that domestication occurred in Central America because of the high regional diversity. Others argued for an origin in northern South America (Giacometti and Leon, 1994; Piperno and Pearsall, 1998; Lebot, 2009; Piperno, 2011), with dispersal into Central America. At this point it is impossible to affirm that cocoyams started to be domesticated in the upper Madeira, even though the diverse ecosystems of the area contain Xanthosoma and it is found in local home gardens and swiddens, where local Indians (Kaxinawá) recognize four ethnovarieties named by the generic term Yubi that can be differentiated by color, such as taioba branca and taioba roxa (Bianchini, 2006).
CONCLUSIONS
There is no doubt about the location of the center of domestication of manioc within the upper Madeira River basin, although immediately adjacent areas may also be involved. Likewise, peach palm has its center of domestication here also, although immediately adjacent areas in the upper Ucayali cannot be ruled out. Capsicum baccatum is firmly located in this area, but evidence for C. frutescens is less clear. Important domesticated populations of peanut are also securely located in the area, although the primary domestication of peanut occurred further south. While there are wild populations of annatto and cocoyam in the region, the information available today is insufficient to affirm that they started to be domesticated in the upper Madeira.
ACKNOWLEDGEMENTS
We thank Claudia López, Museu Paraense Emílio Goeldi (MPEG), and Laure Emperaire, Institut de Recherche pour le Développement (IRD), for the invitation to participate in this Dossiê, Barbara Pickersgill, University of Reading, and two anonymous reviewers for useful criticism and suggestions on the manuscript. CRC thanks the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for a research fellowship (n. 303851/2015-5).
REFERENCES
- ALBRECHT, Elena; ZHANG, Dapeng; SAFTNER, Robert A.; STOMMEL, John R. Genetic diversity and population structure of Capsicum baccatum genetic resources. Genetic Resources and Crop Evolution, Dordrecht, v. 59, n. 4, p. 517-538, 2012.
- ALLEM, Antonio C. The origin and taxonomy of cassava. In: HILLOCKS, Rory J.; THRESH, J. M.; BELLOTTI, Anthony C. (Eds.). Cassava: biology, production and utilization. Oxford: CAB International, 2002. p. 1-16.
- APARICIO, Miguel. Espíritos não humanos, espíritos desumanos: o mundo da sobrenatureza nos Suruwaha do Rio Purus. Espaço Ameríndio, Porto Alegre, v. 9, n. 3, p. 63-85, 2015.
- ARROYO-KALIN, Manuel. The Amazonian Formative: crop domestication and anthropogenic soils. Diversity, Basel, v. 2, n. 4, p. 473-504, 2010.
- BALÉE, William. Antiquity of traditional ethnobiological knowledge in Amazonia: the Tupi-Guarani family and time. Ethnohistory, Durham, v. 47, n. 2, p. 399-422, 2000.
- BARBIERI, Rosa Lía; NEITZKE, R. S. Pimentas do gênero Capsicum. Cor, fogo e sabor. In: BARBIERI, Rosa Lía; STUMPF, Elisabeth R. T. (Eds.). Origem e evolução de plantas cultivadas Brasília: EMBRAPA, 2008. p. 728-745.
- BARBOSA, Reinaldo I.; LUZ, Francisco J. F.; NASCIMENTO FILHO, Herundino R.; MADURO, Cice B. Pimentas do gênero Capsicum cultivadas em Roraima, Amazônia brasileira. I. Espécies domesticadas. Acta Amazonica, Manaus, v. 32, n. 2, p. 177-192, 2002.
- BIANCHINI, Paola C. Agroflorestas e agentes agroflorestais indígenas no Acre 2006. 112 f. Dissertation (Masters in Agroecosystems) – Universidade Federal de Santa Catarina, Florianópolis, 2006.
- BILLOTTE, N.; COUVREUR, T.; MARSEILLAC, N.; BROTTIER, P.; PERTHUIS, B.; VALLEJO, M.; NOYER, J.-L.; JACQUEMOUD-COLLET, J.-P.; RISTERUCCI, A.-M.; PINTAUD, J.-C. A new set of microsatellite markers for the peach palm (Bactris gasipaes Kunth): characterization and across-taxa utility within the tribe Cocoeae. Molecular Ecology Notes, Malden, v. 4, p. 580-582, 2004.
- BOWN, Deni. Aroids: plants of the arum family. 2. ed. Portland: Timber Press, 2000.
- BROWN, Cecil H.; CLEMENT, Charles R.; EPPS, Patience; LUEDELING, Eike; WICHMANN, Soeren. The paleobiolinguistics of domesticated manioc (Manihot esculenta). Ethnobiology Letters, Austin, v. 4, p. 61-70, 2013a.
- BROWN, Cecil H.; CLEMENT, Charles R.; EPPS, Patience; LUEDELING, Eike; WICHMANN, Soeren. The paleobiolinguistics of domesticated chili pepper (Capsicum spp.). Ethnobiology Letters, Austin, v. 4, p. 1-11, 2013b.
- BROWN, Cecil H. Development of agriculture in prehistoric Mesoamerica: the linguistic evidence. In: STALLER, John E.; CARRASCO, Michael (Eds.). Pre-Columbian foodways: interdisciplinary approaches to food, culture, and markets in ancient Mesoamerica. New York: Springer, 2010. p. 71-107.
- CLEMENT, Charles R. 1492 and the loss of Amazonian crop genetic resources. II. Crop biogeography at contact. Economic Botany, New York, v. 53, n. 2, p. 203-216, 1999.
- CLEMENT, Charles R. Pejibaye (Bactris gasipaes). In: SMARTT, Joe; SIMMONDS, Normand W. (Eds.). Evolution of crop plants 2. ed. London: Longman, 1995. p. 383-388.
- CLEMENT, Charles R. Domestication of the pejibaye palm (Bactris gasipaes): past and present. In: BALICK, Michael J. (Ed.) The palm: tree of life. Biology, utilization and conservation. Bronx, New York: The New York Botanical Garden, 1988. (Advances in Economic Botany, v. 6). p. 155-174.
- CLEMENT, Charles R.; DENEVAN, William M.; HECKENBERGER, Michael J.; JUNQUEIRA, André Braga; NEVES, Eduardo G.; TEIXEIRA, Wenceslau G.; WOODS, William I. The domestication of Amazonia before European conquest. Proceedings of the Royal Society B – Biological Sciences, London, v. 282, n. 1812, 20150813, 2015.
- CLEMENT, Charles R.; CRISTO-ARAÚJO, Michelly; D’EECKENBRUGGE, Geo Coppens; ALVES PEREIRA, Alessandro; PICANÇO-RODRIGUES, Doriane. Origin and domestication of native Amazonian crops. Diversity, Basil, v.2, n. 1, p. 72-106, 2010.
- CLEMENT, Charles R.; RIVAL, Laura; COLE, David M. Domestication of peach palm (Bactris gasipaes Kunth): the roles of human mobility and migration. In: ALEXIADES, Miguel N. (Ed.) Shifting spaces, changing times: mobility, migration and displacement in indigenous lowland South America. Oxford: Berghahn Books, 2009a. p. 117-140.
- CLEMENT, Charles R.; SANTOS, Ronaldo P.; DESMOULIERE, Sylvain J. M.; FERREIRA, Evandro J. L.; FARIAS NETO, João T. Ecological adaptation of wild peach palm, its in situ conservation and deforestation-mediated extinction in southern Brazilian Amazonia. PLoS ONE, San Francisco, v. 4, n. 2, e4564, 2009b.
- COLUNGA-GARCÍAMARÍN, Patricia; ZIZUMBO-VILLARREAL, Daniel. Domestication of plants in Maya lowlands. Economic Botany, New York, v. 58, sup. 1, p. S101-S110, 2004.
- CRISTO-ARAÚJO, Michelly; PICANÇO-RODRIGUES, Doriane; ASTOLFI FILHO, Spartaco; CLEMENT, Charles R. Peach palm core collection in Brazilian Amazonia. Crop Breeding and Applied Biotechnology, Viçosa, v. 15, n. 1, p. 18-25, 2015.
- CRISTO-ARAÚJO, Michelly; PICANÇO-RODRIGUES, Doriane; ASTOLFI-FILHO, Spartaco; CLEMENT, Charles R. Genetic variability in the peach palm genebank with RAPD markers. Crop Breeding and Applied Biotechnology, Viçosa, v. 10, n. 3, p. 211-217, 2010.
- CRISTO-ARAÚJO, Michelly; REIS, Vanessa M.; PICANÇO-RODRIGUES, Doriane; CLEMENT, Charles R. Domestication of peach palm in southwestern Amazonia. Tipití, Philadelphia, v. 2, p. 74-80, 2013.
- DICKAU, Ruth; BRUNO, Maria C.; IRIARTE, José; PRÜMERS, Heiko; JAIMES BETANCOURT, Carla; HOLST, Irene; MAYLE, Francis E. Diversity of cultivars and other plant resources used at habitation sites in the Llanos de Mojos, Beni, Bolivia: evidence from macrobotanical remains, starch grains, and phytoliths. Journal of Archaeological Science, Amsterdam, v. 39, n. 2, p. 357-370, 2012.
- DONKIN, Robin A. Bixa orellana: the eternal shrub. Anthropos, Vienna, v. 69, p. 33-56, 1974.
- DUCKE, Adolfo. Plantas de cultura precolombiana na Amazônia brasileira: notas sobre as espécies ou formas espontâneas que supostamente lhes teriam dado origem. Boletim Técnico do Instituto Agronômico do Norte, Belém, v. 8, p. 1-24, 1946.
- ELIAS, Marianne; MÜHLEN, Gilda S.; McKEY, Doyle; ROA, Ana Carolina; TOHME, Joe. Genetic diversity of traditional South American landraces of cassava (Manihot esculenta Crantz): an analysis using microsatellites. Economic Botany, New York, v. 58, n. 2, p. 242-256, 2004.
- ERICKSON, Clark L. Archaeological methods for the study of ancient landscapes of the Llanos de Mojos in the Bolivian Amazon. In: STAHL, Peter W. (Ed.). Archaeology in the lowland American tropics: current analytical methods and applications. New York: Columbia University Press, 1995. p. 66-96.
- ERIKSEN, Love. Nature and culture in prehistoric Amazonia: using GIS to reconstruct ancient ethnogenetic processes from archaeology, linguistics, geography, and ethnohistory. Lund, Sweden: Lund University, 2011.
- ERIKSEN, Love; DANIELSEN, Swintha. The Arawakan matrix. In: O’CONNOR, Loretta; MUYSKEN, Pieter (Eds.). The native languages of South America: origins, development, typology. Cambridge: Cambridge University Press, 2014. p. 152-176.
- ERIKSEN, Love; GALUCIO, Ana Vilacy. The Tupian expansion. In: O’CONNOR, Loretta; MUYSKEN, Pieter (Eds.). The native languages of South America: origins, development, typology. Cambridge: Cambridge University Press, 2014. p. 177-199.
- FÁVERO, Alessandra P.; VALLS, José Francisco M. Domesticação e melhoramento de amendoim. In: BORÉM, Aloízio; LOPES, Maria Teresa G.; CLEMENT, Charles R. (Eds.). Domesticação e melhoramento: espécies amazônicas. Viçosa: Editora da Universidade Federal de Viçosa, 2009. p. 237-249.
- FERREIRA, Evandro. The phylogeny of pupunha (Bactris gasipaes Kunth, Palmae) and allied species. In: HENDERSON, Andrew; BORCHSENIUS, Finn (Eds.). Evolution, variation, and classification of palms New York: The New York Botanical Garden, 1999. (Memoirs of the New York Botanical Garden, v. 83). p. 225-236.
- GERSTEL, D. U.; SISSON, V. A. Tobacco, Nicotiana tabacum (Solanaceae). In: SMARTT, Joe; SIMMONDS, Norman W. (Eds.). Evolution of crop plants 2. ed. London: Longman, 1995. p. 458-463.
- GIACOMETTI, Dalmo C.; LEON, Jorge. Yannia, yautia (Xanthosoma sagittifolium). In: HERNÁNDEZ-BERMEJO, J. E.; LEON, J. (Eds.). Neglected crops: 1492 from a different perspective. Rome: FAO, 1994. (FAO Plant Protection Series, n. 26). p. 255-258.
- GRABIELE, Marina; CHALUP, Laura; ROBLEDO, Germán; SEIJO, Guillermo. Genetic and geographic origin of domesticated peanut as evidenced by 5S rDNA and chloroplast DNA sequences. Plant Systematics and Evolution, Vienna, v. 298, n. 6, p. 1151-1165, 2012.
- HAWKES, Jack G. The domestication of roots and tubers in the American tropics. In: HARRIS, David R.; HILLMAN, Gordon C. (Eds.). Foraging and farming: the evolution of plant exploitation. London: Unwin Hyman, 1989. p. 481-503.
- HEISER JR., Charles B. Peppers. Capsicum (Solanaceae). In: SMARTT, Joe; SIMMONDS, Normand W. (Eds.). Evolution of crop plants 2. ed. London: Longman, 1995. p. 449-451.
- HENDERSON, Andrew. Bactris (Palmae) New York: The New York Botanical Garden, 2000. (Flora Neotropica, v. 79). 181 p.
- HERNÁNDEZ-UGALDE, José A. Descripción de la diversidad y estructura genética de las poblaciones silvestres y cultivadas de pejibaye (Bactris gasipaes Kunth), utilizando marcadores microsatélites 2005. 79 f. Dissertation (Masters in Biology) – Universidad de Costa Rica, San José, 2005.
- HERNÁNDEZ-UGALDE, José A.; MORA-URPÍ, Jorge; ROCHA, Oscar J. Genetic relationships among wild and cultivated populations of peach palm (Bactris gasipaes Kunth, Palmae): evidence for multiple independent domestication events. Genetic Resources and Crop Evolution, Dordrecht, v. 58, n. 4, p. 571-583, 2011.
- HERNÁNDEZ-UGALDE, José A.; MORA URPÍ, Jorge; ROCHANUÑEZ, Oscar. Diversidad genética y relaciones de parentesco de las poblaciones silvestres y cultivadas de pejibaye (Bactris gasipaes, Palmae), utilizando marcadores microsatelites. Revista de Biología Tropical, San José, v. 56, n. 1, p. 217-245, 2008.
- ISENDAHL, Christian. The domestication and early spread of manioc (Manihot esculenta Crantz): a brief synthesis. Latin American Antiquity, Washington, v. 22, n. 4, p. 452-468, 2011.
- JACOBSON, M.; BORNSTEIN, S.; WALLGREN, P.; SCHNELL, R. J.; GOENAGA, R.; OLANO, C. T. Genetic similarities among cocoyam cultivars based on randomly amplified polymorphic DNA (RAPD) analysis. Scientia Horticulturae, Amsterdam, v. 80, n. 3-4, p. 267-276, 1999.
- KRAPOVICKAS, Antonio. El origen y dispersión de las variedades del maní. Academia Nacional de Agronomia y Veterinária, Buenos Aires, v. 49, p. 18-26, 1995.
- KRAPOVICKAS, Antonio; GREGORY, Walton C. Taxonomia del género Arachis (Leguminosae). Bonplandia, Corrientes, v. 8, n. 1-4, p. 1-186, 1994.
- KUNTZE, Carl. Bixaceae. In: ENGLER, A.; PRANTL, K. (Eds.) Die natürlichen flanzenfamilien Leipzig, Germany: Engelmann, 1925. p. 315.
- LAGROU, Els. Rir do poder e o poder do riso nas narrativas e performances Kaxinawa. Revista de Antropologia, São Paulo, v. 49, n. 1, p. 55-90, 2006.
- LAHAYE, Christelle; HERNANDEZ, Marion; BOËDA, Eric; FELICE, Gisele D.; GUIDON, Niède; HOELTZ, Sirlei; LOURDEAU, Antoine; PAGLI, Marina; PESSIS, Anne-Marie; RASSE, Michel; VIANA, Sibeli. Human occupation in South America by 20,000 BC: the Toca da Tira Peia site, Piauí, Brazil. Journal of Archaeological Science, Amsterdam, v. 40, n. 6, p. 2840-2847, 2013.
- LEBOT, Vincent. Tropical root and tuber crops: cassava, sweet potato, yams and aroids. Oxford: CAB International, 2009.
- LEÓN, Jorge. Botánica de los cultivos tropicales, 2 ed. San José, Costa Rica: Editorial Agroamérica, Instituto Interamericano de Cooperación para la Agricultura, 2000.
- LÉVI-STRAUSS, Claude. Mitologicas II - do mel às cinzas São Paulo: Cosac e Naify, 2004.
- LÉVI-STRAUSS, Claude. The use of wild plants in tropical South America. Economic Botany, New York, v. 6, p. 252-270, 1957.
- LÉVI-STRAUSS, Claude. The Nambicuara. In: STEWARD, Julian (Ed.). Handbook of South American Indians Washington: Smithsonian Institution Press, 1948. p. 371-379.
- LOMBARDO, Umberto; SZABO, Katherine; CAPRILES, Jose M.; MAY, Jan-Hendrik; AMELUNG, Wulf; HUTTERER, Rainer; LEHNDORFF, Eva; PLOTZKI, Anna; VEIT, Heinz. Early and middle holocene hunter-gatherer occupations in western Amazonia: the hidden shell middens. PLoS ONE, San Francisco, v. 8, n. 8, e72746, 2013.
- MARTÍNEZ, A. K.; GAITÁN-SOLIS, E.; DUQUE, M. C.; BERNAL, R.; TOHME, J. Microsatellite loci in Bactris gasipaes (Arecaceae): their isolation and characterization. Molecular Ecology Notes, Malden, v. 2, p. 408-410, 2002.
- MARTINS, Matheus; TEIXEIRA, Marcos. Cenas da vida do Amazonas: um estudo dos contos de Inglês de Sousa. Revista de Literatura, Belo Horizonte, p. 31-62, 2005.
- McKEY, Doyle; BECKERMAN, Stephen. Chemical ecology, plant evolution and traditional manioc cultivation systems. In: HLADIK, Claude M.; HLADIK, Annette; LINARES, Olga F.; PAGEZY, Hélène; SEMPLE, Alison; HADLEY, Malcolm (Eds.). Tropical forests, people and food: biocultural interactions and applications to development. Carnforth: Parthenon; Paris: UNESCO, 1993. p. 83-112
- MEYER, Rachel S.; PURUGGANAN, Michael D. Evolution of crop species: genetics of domestication and diversification. Nature Reviews Genetics, London, v. 14, n. 12, p. 840-852, 2013.
- MILLER, Eurico T. Adaptação agrícola pré-histórica no alto rio Madeira. In: MEGGERS, Betty J. (Ed.). Prehistoria sudamericana: nuevas perspectivas. Washington: Taraxacum, 1992a. p. 219-232.
- MILLER, Eurico T. (Coord.). Arqueologia nos empreendimentos hidrelétricos da Eletronorte: resultados preliminares. Brasília: Eletronorte, 1992b. 93 p.
- MORA URPÍ, Jorge. Origen y domesticación. In: MORA-URPÍ, Jorge; GAINZA ECHEVERRÍA, Javier (Eds.). Palmito de Pejibaye (Bactris gasipaes Kunth): su cultivo e industrialización. San José, Costa Rica: Editorial de la Universidad de Costa Rica, 1999. p. 17-24.
- MORA URPÍ, Jorge; WEBER, John C.; CLEMENT, Charles R. Peach palm Bactris gasipaes Kunth. Promoting the conservation and use of underutilized and neglected crops, Gatersleben, Germany: Institute of Plant Genetics and Crop Plant Research (IPK); Rome: International Plant Genetic Resources Institute (IPGRI), 1997. v. 20.
- MORCOTE-RIOS, Gaspar; BERNAL, Rodrigo. Remains of palms (Palmae) at archaeological sites in the New World: a review. The Botanical Review, New York, v. 67, n. 3, p. 309-350, 2001.
- MOREIRA, Priscila A.; LINS, Juliana; DEQUIGIOVANNI, Gabriel; VEASEY, Elizabeth A.; CLEMENT, Charles R. The domestication of annatto (Bixa orellana) from Bixa urucurana in Amazonia. Economic Botany, New York, v. 69, n. 2, p. 127-135, 2015.
- MÜHLEN, GildaS.; ALVESPEREIRA, Alessandro.; CLEMENT, CharlesR.; VALLE, TeresaLosada. Genetic diversity and differentiation of Brazilian bitter and sweet manioc varieties (Manihot esculenta Crantz, Euphorbiaceae) based on SSR molecular markers. Tipití, Philadelphia, v. 11, n. 2, p. 66-73, 2013.
- MÜHLEN, Gilda S.; MARTINS, Paulo S.; ANDO, Akihiko. Variabilidade genética de etnovariedades de mandioca, avaliada por marcadores de DNA. Scientia Agricola, Piracicaba, v. 57, n. 2, p. 319-328, 2000.
- NEI, Masatoshi. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics, Bethesda, v. 23, p. 341-369, 1978.
- NEVES, Eduardo G.; PUGLIESE JR.; Francisco A.; SHOCK, Myrtle Pearl; ZIMPEL NETO, Carlos A.; HERMENEGILDO, Tiago. Mapping and preliminary excavations at the Monte Castelo shell mound, Southwestern Amazonia: report submitted to the Committee on Exploration, National Geographic Society. São Paulo: Laboratório de Arqueologia dos Trópicos/Museu de Arqueologia e Etnologia/Universidade de São Paulo, 2015. 30 p.
- NORTON, Marcy. Sacred gifts, profane pleasures A history of tobacco and chocolate in the Atlantic World. Ithaca: Cornell University Press, 2008.
- OCAMPO, Rafael A. S. Aspectos agronomicos sobre el cultivo del achiote (Bixa orellana L.) In: ARCE P., Jorge (Ed.). Aspectos sobre el achiote y perspectivas para Costa Rica Turrialba: CATIE, 1983. (Informe Técnico, 47). p. 43-57.
- OLSEN, Kenneth M. SNPs, SSRs and inferences on cassava’s origin. Plant Molecular Biology, New York, v. 56, n. 4, p. 517-526, 2004.
- OLSEN, Kenneth M.; SCHAAL, Barbara A. Evidence on the origin of cassava: phylogeography of Manihot esculenta Proceedings of the National Academy of Sciences of the United States of America, Washington, v. 96, n. 10, p. 5586-5591, 1999.
- OLSEN, Kenneth M.; SCHAAL, Barbara A. Microsatellite variation in cassava (Manihot esculenta, Euphorbiaceae) and its wild relatives: further evidence for a southern Amazonian origin of domestication. American Journal of Botany, Saint Louis, v. 88, n. 1, p. 131-142, 2001.
- PEARSALL, Deborah M. The origins of plant cultivation in South America. In: COWAN, C. Wesley; WATSON, Patty Jo (Eds.). The origins of agriculture: an international perspective. Washington: Smithsonian Institution Press, 1992. p. 173-206.
- PERRUT-LIMA, Poliana; MÜHLEN, Gilda S.; CARVALHO, Cassia R. L. Cyanogenic glycoside content of Manihot esculenta subsp. flabellifolia in south-central Rondônia, Brazil, in the center of domestication of M. esculenta subsp. esculenta Genetic Resources and Crop Evolution, Dordrecht, v. 61, n. 6, p. 1035-1038, 2014.
- PICKERSGILL, Barbara. Domestication of plants in the Americas: Insights from Mendelian and molecular genetics. Annals of Botany, Oxford, v. 100, n. 5, p. 925-940, 2007.
- PICKERSGILL, Barbara. The domestication of Chili peppers. In: UCKO, Peter J.; DIMBLEBY, G. W. (Ed.). The domestication and exploitation of plants and animals London: Duckworth, 1969. p. 443-450.
- PIPERNO, Dolores R. The origins of plant cultivation and domestication in the New World tropics: patterns, process, and new developments. Current Anthropology, Chicago, v. 52, n. S4, p. S453-S470, 2011.
- PIPERNO, Dolores R.; PEARSALL, Deborah M. The origins of agriculture in the Lowland Neotropics San Diego: Academic Press, 1998.
- PLOWMAN, Timothy. The origin, evolution, and diffusion of coca, Erythroxylum spp. in South and Central America. In: STONE, Doris (Ed.). Pre-Columbian plant migration Cambridge: Harvard University, 1984. (Papers of the Peabody Museum of Archaeology and Ethnology, v. 76), p. 125-164.
- PLUCKNET, Donald L. Edible aroids. In: SIMMONDS, Normand W. (Ed.). Evolution of crop plants London: Longman, 1976. p. 10-12.
- PRITCHARD, Jonathan K.; STEPHENS, Mathew; DONNELLY, Peter. Inference of population structure using multilocus genotype data. Genetics, Bethesda, v. 155, n. 2, p. 945-959, 2000.
- PURSEGLOVE, John W. Tropical crops: monocotyledons. London: Longman, 1972.
- REIFSCHNEIDER, Francisco J. B. Capsicum: pimentas e pimentões no Brasil. Brasília: EMBRAPA Comunicação para Transferência de Tecnologia, 2000.
- REIS, Vanessa M. Relações genéticas entre raças e populações da Coleção Nuclear de Pupunha (Bactris gasipaes Kunth) avaliadas com microssatélites 2009. 86 f. Dissertation (Masters in Tropical Agronomy) – Universidade Federal do Amazonas, Manaus, 2009.
- RODRIGUES, Doriane P.; ASTOLFI FILHO, Spartaco; CLEMENT, Charles R. Molecular marker-mediated validation of morphologically defined landraces of pejibaye (Bactris gasipaes) and their phylogenetic relationships. Genetic Resources and Crop Evolution, Dordrecht, v. 51, n. 8, p. 871-882, 2004a.
- RODRIGUES, Doriane P.; VINSON, Cristina; CIAMPI, Anna Y.; FARIAS, Izeni P.; LEMES, Maristerra R.; ASTOLFI-FILHO, Spartaco; CLEMENT, Charles R. Novel microsatellite markers for Bactris gasipaes (Palmae). Molecular Ecology Notes, Malden, v. 4, n. 4, p. 575-576, 2004b.
- ROGERS, David J.; APPAN, Subramaniam G. Manihot and Manihotoides (Euphorbiaceae): a computer-assisted study. New York: Hafner Press, 1973. (Flora Neotropica, Monograph n. 13).
- ROOSEVELT, Anna C. The Amazon and the Anthropocene: 13,000 years of human influence in a tropical rainforest. Anthropocene, Amsterdam, v. 4, p. 69-87, 2013.
- SAUER, Carl O. As plantas cultivadas na América do Sul tropical. In: RIBEIRO, Darcy (Ed.). Suma etnobiológica brasileira: etnobiologia. 2. ed. Petrópolis: Vozes, 1987. p. 151-158.
- SCHAAL, Barbara A.; OLSEN, Ken M.; CARVALHO, Luis J. C. B. Evolution, domestication, and agrobiodiversity in the tropical crop cassava. In: MOTLEY, Timothy J.; ZEREGA, Nyree; CROSS, Hugh (Eds.). Darwin’s harvest: new approaches to the origins, evolution, and conservation of crops. New York: Columbia University Press, 2006. p. 269-284.
- SCHULTES, Richard E. Amazonian cultigens and their northward and westward migrations in pre-Columbian times. In: STONE, Doris (Ed.). Pre-Columbian plant migration Cambridge: Harvard University, 1984. (Papers of the Peabody Museum of Archaeology and Ethnology, v. 76). p. 19-38.
- SHAW, Joey; LICKEY, Edgar B.; SCHILLING, Edward E.; SMALL, Randall L. Comparison of whole chloroplast genome sequences to choose noncoding regions for phylogenetic studies in angiosperms: the tortoise and the hare. III. American Journal of Botany, Saint Louis, v. 94, n. 3, p. 275-288, 2007.
- SILVA, Andréa L. Comida de gente: preferências e tabus alimentares entre os ribeirinhos do Médio Rio Negro (Amazonas, Brasil). Revista de Antropologia, São Paulo, v. 50, n. 1, p. 125-179, 2007.
- VALLS, José Francisco M.; SIMPSON, Charles E. New species of Arachis (Leguminosae) from Brazil, Paraguay and Bolivia. Bonplandia, Corrientes, v. 14, n. 1-2, p. 35-63, 2005.
- WALKER, Robert S.; RIBEIRO, Lincoln A. Bayesian phylogeography of the Arawak expansion in lowland South America. Proceedings of the Royal Society B - Biological Sciences, London, v. 278, n. 1718, p. 2562-2567, 2011.
- WATLING, Jennifer; SAUNALUOMA, Sanna; PÄRSSINEN, Martti; SCHAAN, Denise. Subsistence practices among earthwork builders: phytolith evidence from archaeological sites in the southwest Amazonian interfluves. Journal of Archaeological Science: Reports, Amsterdam, v. 4, p. 541-551, 2015.
- WHITNEY, Bronwen S.; DICKAU, Ruth; MAYLE, Francis E.; WALKER, John H.; SOTO, J. Daniel; IRIARTE, José. Pre-Columbian raised-field agriculture and land use in the Bolivian Amazon. The Holocene, Los Angeles, v. 24, n. 2, p. 231-241, 2014.