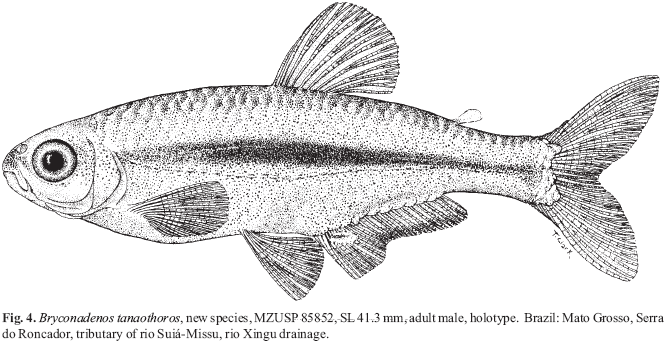
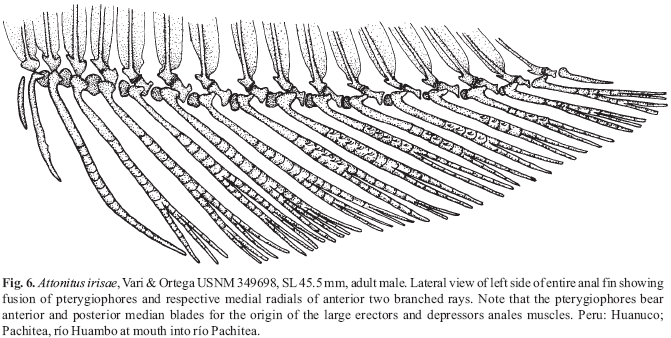
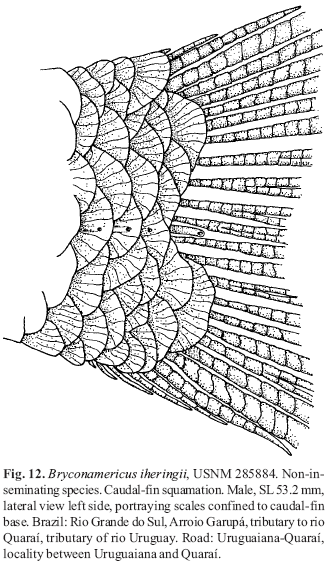
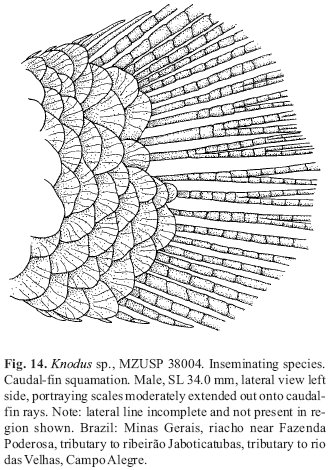
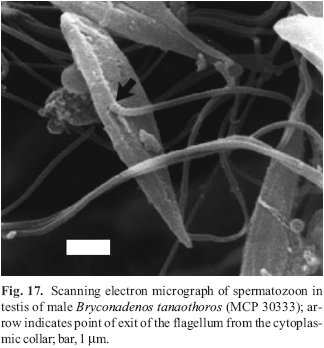
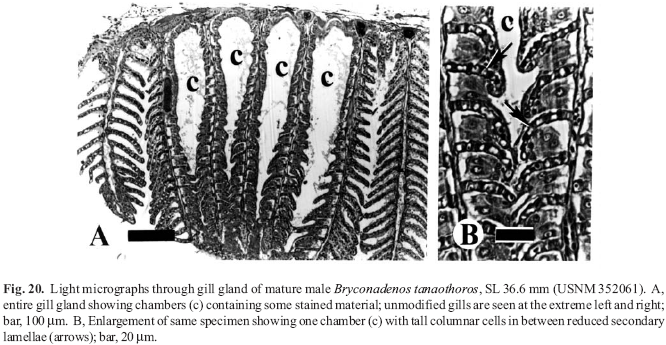
A new inseminating fish species of the family Characidae, Bryconadenos tanaothoros, from tributaries of the upper rio Xingu and upper rio Tapajós basins, Mato Grosso, Brazil is described as the type species of a new genus. This new species and the genus are characterized by a glandular organ on the anterior region of the anal fin of sexually mature males, curved lower jaw teeth, and an inseminating reproductive mode. This new genus is hypothesized as most closely related to Attonitus, a genus with three inseminating species from Peru. Bryconadenos and Attonitus are suggested as related to certain inseminating, but undescribed characid species of uncertain relationships that are similar in certain respects to species of the glandulocaudine Planaltina and to the inseminating species of Knodus. These and a few other inseminating characids are included in a previous tentative characid subgroup designated as Clade A. No species among a relatively small sample of the many species of the Clade A genus Bryconamericus were found inseminating, except Bryconamericus pectinatus. However, newly collected specimens of B. pectinatus were found to have caudal-fin squamation like that of the species of Knodus and this species is here tentatively referred to Knodus. Our investigations indicate that at least several species of Knodus, including the type species, Knodus meridae, are not inseminating, but we found two inseminating apparently new characid species that currently would be referred to Knodus. These species lack the derived anal-fin rays present in the males of K. pectinatus. Other Clade A taxa known to be inseminating, such as two species of the large genus Creagrutus, three species of Monotocheirodon (two undescribed), and the species and genera of the characid subfamily Glandulocaudinae are briefly discussed regarding possible relationships to Attonitus and Bryconadenos. The anatomical aspects of the primary and secondary sexual characteristics of Bryconadenos and Attonitus are discussed in relation to certain other inseminating characids, such as the species of Brittanichthys and Hollandichthys, that are not currently hypothesized to belong to Clade A and presumably acquired insemination independently. It is concluded that much additional data regarding the reproductive modes as well as other anatomical/physiological systems of characids currently included in and excluded from Clade A are necessary before a reasonably supported phylogeny of Clade A characids and their possible outgroup relatives can be advanced. The anal-fin gland cells of sexually active male Bryconadenos specimens are histologically indistinguishable from club cells (also called alarm substance cells) found within the skin of cypriniforms, characiforms, catfishes, and other otophysan fishes. These cells occur at the skin's surface of the anal-fin gland in male Bryconadenos where they are organized into an organ. Many other adult male characids have club cells at the anal-fin's skin surface, often associated with anal-fin hooks, but were not found organized into an organ as in Bryconadenos. We hypothesize these cells to secrete a pheromone during courtship via holocrine secretion. Males of the genera Lophiobrycon , Glandulocauda, and Mimagoniates, tribe Glandulocaudini, were found to have club cells associated with their caudal-fin organ, but no specialized mucus cells were present as found in the caudal organ of males of the glandulocaudine Corynopoma riisei, tribe Stevardiini (= Corynopomini of past authors). In this species, males have hypertrophied mucus cells hypothesized to be modified for pheromone secretion. Evidence that the derived scales and fin rays of the caudal organ of males of the tribe Glandulocaudini are not homologous with that of other tribes of the Glandulocaudinae, as this subfamily was previously recognized, is discussed and it is concluded that the members of the tribe Glandulocaudini should be recognized as a separate subfamily, the Glandulocaudinae, with possible close relationships to some other Clade A inseminating characids that lack caudal-fin organs. The remaining tribes of the former Glandulocaudinae are here included in the subfamily name Stevardiinae. Many species of these two subfamilies and some of the other inseminating Clade A characids have modified sperm cells with an elongate "binding"cytoplasmic collar and mitochondria located along and beyond the nucleus. This may be an indication of a relationship at a unique level within Clade A characids. However, further research on the derived nature and homology of the sperm cells within inseminating Clade A characids and of the caudal organs of the tribes of the Stevardiinae must be undertaken in order to utilize sperm cell features as characters for studying phylogeny. Finally, the kinds of secretory cells and gross anatomical structures in the tail organs of the stevardiine tribes need detailed research in order to better present hypotheses of phylogeny for the tribes of the Stevardiinae as well as all inseminating Clade A characids.
Inseminating characid phylogeny; putative male pheromone cells; Attonitus; Bryconadenos; Knodus