Abstracts
The present study compares the reproduction, condition and size of the small characiform fish, Characidium pterostictum, sampled at close sites differing in severity of flash flood effects. Data were obtained from seasonal samples in two sites situated 8 km apart in the same stream. In the upstream site, habitat is more severely affected by flash floods than in the downstream site, and this difference was hypothesized to produce differences in life history and individual reproduction trade-off patterns, as predicted by life-history theory. The results provided evidence for small-scale spatial variation in life-history and trade-off patterns within the studied population. At the most severely disturbed site, C. pterostictum displayed a trade-off pattern that favored reproductive life-span (e.g., larger size, higher and seasonally stable condition, larger mean size of mature females) over instantaneous reproductive output (lower gonadosomatic index), while the opposite pattern was observed in the less disturbed site. Because of the differences in disturbance effects between each sampling site, these results suggest that within-stream variability in the severity of hydrological disturbance can influence life-history patterns at small spatial scales. An implication of the results is that fish occupying areas that are hydrologically more variable within a stream are not necessarily at an energetic or reproductive disadvantage, but may be simply under environmental conditions that favor distinct patterns of energy allocation (or trade-offs) and population persistence, as predicted by life-history theory. Therefore, plasticity in life-history is expected to be common in stream fish populations that are widespread in a stream system with spatially variable or patchy habitat characteristics.
Fish; Condition; Reproductive trade-off; Natural disturbance
Este estudo compara a estrutura populacional e a reprodução do pequeno caraciforme, Characidium pterostictum, amostrado em locais sujeitos a diferentes efeitos de perturbações hidrológicas naturais (enxurradas). Os dados foram obtidos a partir de amostragens sazonais em dois trechos distando 8 km entre si, no mesmo riacho. No trecho a montante, o habitat é mais severamente afetado pelas enxurradas do que no trecho a jusante e, devido a esta diferença, testou-se a hipótese de que haveria contrastes em padrões de alocação de energia entre reprodução e sobrevivência individual, como previsto pela teoria de história de vida. No trecho a montante, C. pterostictum apresentou características que favorecem a longevidade reprodutiva (como maior tamanho, condição mais elevada e sazonalmente pouco variável, maior tamanho de fêmeas maduras) em relação ao desempenho reprodutivo instantâneo (menor índice gonadossomático). O padrão oposto foi observado no trecho a jusante. Devido às diferenças em termos de efeito de perturbação (enxurradas) em cada trecho, esses resultados sugerem que a variabilidade espacial de perturbações hidrológicas pode influenciar padrões de história de vida dentro de uma população. Outra implicação dos resultados, é que os peixes que ocupam locais hidrologicamente mais variáveis em um riacho não estão necessariamente em desvantagem energética ou reprodutiva, mas podem estar simplesmente sob condições ambientais que favorecem padrões distintos de alocação de energia e persistência da população. Dessa forma, pode-se esperar que a plasticidade de história de vida seja comum em populações de peixes que sejam amplamente distribuídas em um sistema de riachos com características de hábitat espacialmente variáveis ou em mosaico.
Life-history of the South American darter, Characidium pterostictum (Crenuchidae): evidence for small scale spatial variation in a piedmont stream
Fernando Gertum Becker; Silene de Carvalho; Sandra Maria Hartz
Departamento de Ecologia, Universidade Federal do Rio Grande do Sul, Av. Bento Gonçalves, 9500, Caixa Postal 15007, 91501-970 Porto Alegre, RS, Brazil. sandra.hartz@ufrgs.br
ABSTRACT
The present study compares the reproduction, condition and size of the small characiform fish, Characidium pterostictum, sampled at close sites differing in severity of flash flood effects. Data were obtained from seasonal samples in two sites situated 8 km apart in the same stream. In the upstream site, habitat is more severely affected by flash floods than in the downstream site, and this difference was hypothesized to produce differences in life history and individual reproduction trade-off patterns, as predicted by life-history theory. The results provided evidence for small-scale spatial variation in life-history and trade-off patterns within the studied population. At the most severely disturbed site, C. pterostictum displayed a trade-off pattern that favored reproductive life-span (e.g., larger size, higher and seasonally stable condition, larger mean size of mature females) over instantaneous reproductive output (lower gonadosomatic index), while the opposite pattern was observed in the less disturbed site. Because of the differences in disturbance effects between each sampling site, these results suggest that within-stream variability in the severity of hydrological disturbance can influence life-history patterns at small spatial scales. An implication of the results is that fish occupying areas that are hydrologically more variable within a stream are not necessarily at an energetic or reproductive disadvantage, but may be simply under environmental conditions that favor distinct patterns of energy allocation (or trade-offs) and population persistence, as predicted by life-history theory. Therefore, plasticity in life-history is expected to be common in stream fish populations that are widespread in a stream system with spatially variable or patchy habitat characteristics.
Key words: Fish, Condition, Reproductive trade-off, Natural disturbance.
RESUMO
Este estudo compara a estrutura populacional e a reprodução do pequeno caraciforme, Characidium pterostictum, amostrado em locais sujeitos a diferentes efeitos de perturbações hidrológicas naturais (enxurradas). Os dados foram obtidos a partir de amostragens sazonais em dois trechos distando 8 km entre si, no mesmo riacho. No trecho a montante, o habitat é mais severamente afetado pelas enxurradas do que no trecho a jusante e, devido a esta diferença, testou-se a hipótese de que haveria contrastes em padrões de alocação de energia entre reprodução e sobrevivência individual, como previsto pela teoria de história de vida. No trecho a montante, C. pterostictum apresentou características que favorecem a longevidade reprodutiva (como maior tamanho, condição mais elevada e sazonalmente pouco variável, maior tamanho de fêmeas maduras) em relação ao desempenho reprodutivo instantâneo (menor índice gonadossomático). O padrão oposto foi observado no trecho a jusante. Devido às diferenças em termos de efeito de perturbação (enxurradas) em cada trecho, esses resultados sugerem que a variabilidade espacial de perturbações hidrológicas pode influenciar padrões de história de vida dentro de uma população. Outra implicação dos resultados, é que os peixes que ocupam locais hidrologicamente mais variáveis em um riacho não estão necessariamente em desvantagem energética ou reprodutiva, mas podem estar simplesmente sob condições ambientais que favorecem padrões distintos de alocação de energia e persistência da população. Dessa forma, pode-se esperar que a plasticidade de história de vida seja comum em populações de peixes que sejam amplamente distribuídas em um sistema de riachos com características de hábitat espacialmente variáveis ou em mosaico.
Introduction
Streams are heterogeneous environments where physical and temporal variability influence biotic patterns and processes, including fish life-history (Poff & Ward, 1990). Environmental variability, particularly discharge variability and natural disturbance (droughts and flash floods), have often been related to fish life-history (e.g. Winemiller, 1989; Vila-Gispert & Moreno-Amich, 2002; Magalhães et al., 2003). In fish populations facing high environmental variability or adverse conditions (e.g., food limitation), there is a trade-off between individual survival and reproduction (Winemiller, 2004), with some species sacrificing egg production for growth, while others sacrifice condition to maintain fecundity (Link & Burnett, 2001).
Most studies, however, have compared life-history between different habitat types, between species, or considered relatively wide temporal and spatial scales, with few cases focusing on small spatial scale variation within populations (Machado et al., 2002; Lobón-Cerviá & Rincón, 2004; Olsen & Volestad, 2003; Olsen & Volestad, 2005). Investigating small-scale spatial variation in life-history is important for understanding how stream fish populations persist under the environmental variability (Magalhães et al., 2003), since mortality is locally variable within streams (Lobón-Cerviá & Rincón, 2004). To cope with local variability in habitat conditions, fish can exhibit different energy allocation patterns among survival, growth, and reproduction (Lobón-Cerviá & Rincón, 2004). A general prediction is that fish inhabiting highly variable habitats are affected by density-independent mortality, and tend to exhibit shorter longevity, smaller body size and earlier maturity, in comparison with less variable environments (Schlosser, 1990; Magalhães et al., 2003; Danylchuk & Tonn, 2006).
The present study describes the life-history of a typical resident stream fish (Characidium pterostictum Gomes, 1947), from piedmont streams of southern Brazil. Small-scale spatial variation in life-history was investigated by comparing fish from the same population, but sampled in two 8-km apart sites that differ in severity of flash flood effects. Life-history trade-off patterns were based on reproductive characteristics (reproductive period, size at maturity, gonadosomatic index), body condition and individual size. We tested the hypothesis that fish in the most severely affected site should exhibit higher investment in present reproduction, with a trade-off for smaller body size and lower condition.
Material and Methods
Study site
Characidium pterostictum was studied in a piedmont stream (Lajeado river; 29º34'S 50º16'W) at the Maquiné river watershed (550 km2), a small Atlantic coastal drainage in southern Brazil. Geology of the Maquiné catchment is mainly basaltic and land cover is a mosaic of agricultural land and regenerating Atlantic Forest (Becker et al., 2004). Climate is subtropical humid, with mean temperatures higher than 22ºC in the warmest month, and between 13-15ºC in the coldest month (July). There is no typical climatic dry season, and rainfall is frequent all over the year (mean annual rainfall is 1400-1800 mm; Hasenack & Ferraro, 1989). Rain storms are common in Summer and result in high energy flash floods that significantly affect the streams in the region.
Lajeado river is the main tributary of the Maquiné river, and has a marked longitudinal profile, flowing from an elevation of 800 m to 200 m a.s.l. in only 5 km. From the headwaters situated in the basaltic plateau, it flows down through the steep canyon-like slope of the Serra Geral, eventually reaching more gentle terrain in the piedmont valleys. In this later section, it has a riffle-pool morphology, with clear water and depths of few centimeters in riffles to about 2.5 m in pools. Pebbles, cobbles and boulders form the stream substrate.
Study species
Characidium pterostictum (Characiformes, Crenuchidae) is one of the most abundant and widely distributed fish species in the streams of the Maquiné river basin (Becker, 2002a). It is commonly found in small streams from southern coastal Atlantic drainages in Brazil, and also in the Uruguay-Paraná basin (Buckup, 2003). Usually, C. pterostictum is found in shallow, clear waters and rocky bottom, being reported to prefer unshaded sites with moderate water flow (Aranha et al., 1998; Bozzetti & Schulz, 2004). Characidium pterostictum occurs almost continuously along the Lajeado stream, with no physical barriers preventing the interchange of individual between the two study sites.
Fish collection and data analysis
Two sites were sampled (PA and PB) in the low-gradient piedmont section of the river. PA is situated 8 km upstream from PB, and has higher gradient and faster water flow over a substrate formed by larger stones (Table 1). During the study period, we observed that PA is more affected by flash floods than PB, since habitat configuration was remarkably modified after severe flash floods in January 2000 (altered size composition of the substrate, macrophyte cover greatly reduced, talweg position altered from mid to left channel). In contrast, habitat conditions in PB remained relatively unchanged after the flash floods.
Individuals of C. pterostictum were collected by kick-sampling in the two riffles, using a rectangular-frame net (frame aperture 45 x 70 cm) with a 1-mm mesh size and a 75-cm deep bag. Samples were taken during the day, in August and November 1999 (Winter and Spring, respectively) and January and May 2000 (Summer and Fall). The stream length sampled varied from 66 to 98 m, depending on water level at the sampling date. The total number of kick-samples varied from 80 to 202, being proportional to the riffle area (see Becker, 2002a, for a description of the sampling design). Collected fish were fixed in 4% formalin and stored in 70% ethanol. Voucher specimens were stored at the Museu de Ciências e Tecnologia of the Pontifícia Universidade Católica do Rio Grande do Sul (MCP 26112, MCP 25355).
Size structure, sex ratio, reproduction and fish condition were used to describe the life history of C. pterostictum and for comparisons between two sampling sites. The total sample comprised 260 specimens (121 males, 129 females, 10 undetermined). In January 2000 (Summer), no specimen was captured in PA, probably because a catastrophic flash flood had occurred only a few days before sampling. From each fish, we took data on total length (Lt; to the nearest mm) and total weight (Wt; 0.01 g), sex, stage of gonadal development (by macroscopic visual examination; Vazzoler, 1996) and gonad weight (Wg; 0.001 g).
The individual size and sex ratio data for samples from each site-date were separately examined. Statistical differences in male: female frequencies were tested by heterogeneity chi-square analysis (Zar, 1999). For each sex, differences in total length between the upstream (PA) and downstream (PB) sites were tested using the Mann-Whitney non-parametric test.
The reproductive period was determined by examining the seasonal variation of the gonadosomatic index, GSI = 100 (Wg .Wt-1), and by analyzing the seasonal frequency of gonadal development stages (non-reproductive, advanced maturing, initial maturing and mature). The "non-reproductive" stage refers to specimens that exhibited reduced gonads and could not be properly assigned to any of the previously mentioned stages. Non-parametric ANOVA (Kruskal-Wallis) and the pair-wise multiple comparison Dunn's test (Zar, 1999) were used to test overall differences in GSI among seven operational sampling units that represented site-season combinations (e.g., PA-Summer, PB-Summer, PA-Fall, PB-Fall, and so on). Size at maturity (Lm) was defined as the length class in which 50% of the specimens are adults. Any individual with gonads in the initial maturing, advanced maturing, and mature stages was considered to be adult. Individuals with "non-reproductive" gonads were not considered adults, although some of these may represent post-reproductive stages with very reduced gonads.
The relative condition factor (Le Cren, 1951) was calculated as K = Wt-g(aLb), where Wt-g = Wt-Wg, and a and b are parameters of the length-weight relationship. The length-weight relationship (Wt = aLtb) was obtained by simple linear regression of natural log-transformed data for females and males (Anderson & Newmann, 1996). The t-test procedures described in Zar (1999) were employed to test for significant differences between males and females and to test for allometry in the length-weight relationship. After testing data for normality and homogeneity of variances, significant differences in condition were tested using a two-way ANOVA, with sampling sites and sampling seasons as factors.
Results
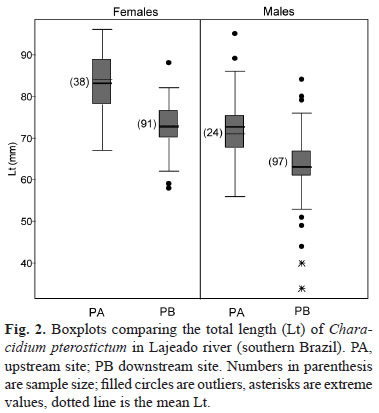
Total length in the samples ranged from 34 mm to 96 mm, with females reaching larger sizes than males (Fig. 1). In PA, specimens were significantly larger than in PB (females, P < 0.001; U' = 418.5; df = 1; males, P < 0.001; U' = 422.5; df = 1) (Fig. 2). Sex ratio for the total sample (129 females, 121 males) was approximately 1:1. However, examination of the seasonal frequencies in each site indicated that sex ratios in PA were different from 1:1 in Winter and Spring (less than 0.5 males:1 female) and Fall (3.3 males:1 female) (heterogeneity χ2 = 9,73; df = 2; P<0.05). In PB, sex ratio remained 1:1 all year round (heterogeneity χ2 = 3,22; df = 3; P<0.25).
A linear relationship between gonad weight and body weight was found for each gonadal development stage, allowing the use of GSI as an indicator of reproductive activity. Gonads of C. pterostictum represented a large proportion of total body weight, with GSI values reaching up to 32.4% in mature females (mean ± sd; 22.2 ± 3.6; n = 45) and 15.1% in mature males (10.0 ± 2.2; n = 44).
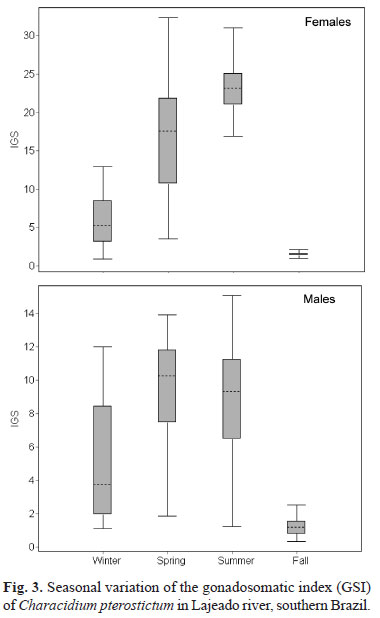
The overall results indicate a long reproductive season for C. pterostictum. Spawning took place between Spring (November) and Summer (February), as shown by the higher GSI of females (Fig. 3) and the larger proportion of mature individuals (Fig. 4). The GSI was significantly different between the upstream (PA) and downstream (PB) sites along seasons of the year for both males (H = 69.9; df = 6; P < 0.001) and females (H = 98.4; df = 6; P < 0.001), with PB usually showing higher values than PA. A recruitment period could not be clearly defined, but smaller specimens were relatively more abundant in Summer and Fall, after the main reproductive period (Spring-Summer). In PA (upstream site), only 2% of the total number of individuals were smaller than 60 mm, while in PB 16.1% were smaller than 60 mm, suggesting that recruitment may have been more effective in PB (Fig. 1). A further evidence for local differences in recruitment was the capture of five immature specimens (undetermined sex; 40 to 60 mm Lt) in PB in the Fall sample.
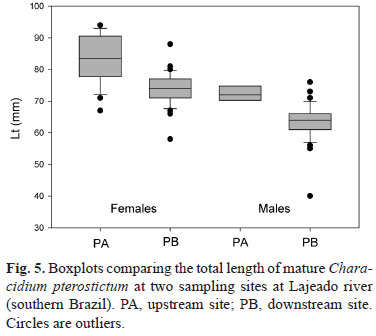
Females tended to reach maturity at larger sizes (Lm = 65-69 mm) than males (Lm = 55-59 mm). The smallest mature female and male specimens occurred respectively in the 50-55 mm in the 45-49 mm length classes. Since the smallest male was the only specimen in its size class, it was ignored in the determination of size at maturity. Length of mature males and females was larger upstream (Fig. 5) (Mann-Whitney test, U = 144; P < 0.001 for females; U = 72; P < 0.001 for males) indicating that upstream individuals tended to reproduce later (assuming equivalent growth rate at both sites).
The length-weight relationship was significantly different between males and females (t = 4.12; df = 230; P<0.05) and the increment of weight relative to length was positive allometric (males and females, P<0.05) (Table 2). The condition of females did not change significantly along the year in PA (F = 0.741; df = 2; P = 0.484), while in PB there was a significant lower condition in Spring and Summer (reproductive season) relative to Fall and Winter (F = 9.698; df = 3; P < 0.001) (Fig. 6). The ANOVA results indicated that significant differences in female condition between the upstream and downstream sites (PA and PB) were dependent on the time of the year, with distinct values observed only in Spring (reproduction), when PB females had the lowest overall condition (t test, t = 2.203; df = 37; P = 0.034). On the other hand, males showed no significant change in condition along the year, both in the upstream (F = 0.512; df = 2; P = 0.606) and the downstream site (H = 5.228; df = 3; P = 0.156; the Kruskal-Wallis test was used because of non-normality in the data), but exhibited statistical differences between the two sites (PA higher than PB; t test on the pooled data of each site; t = 2.919; df = 22; P = 0.008).
A summary of the results (Table 3) provides evidence for several differences in reproductive, condition and size attributes of C. pterostictum between the two study sites. The interpretation of these differences (see the last rows in Table 3) is discussed in the following section.
Discussion
The reproductive period of C. pterostictum is relatively long (4-6 months) and seasonal, with peak reproductive activity in Summer, although part of the population is already spawning in Spring. This pattern is common in most freshwater fish (Matthews, 1998), including those in southern Brazil (e.g., Hartz et al., 1999; Azevedo et al., 2000; Becker, 2002b).
In the study region, apart from temperature and photoperiod, Spring-Summer flash floods are probably the most typically seasonal abiotic factor in piedmont streams. Some species (including one Characidium) from similar environments exhibit an even longer reproductive period than C. pterostictum, what has been hypothesized to be part of a life-history strategy related to the high frequency of natural hydrological disturbances (Mazzoni & Petito, 1999; Spranza & Stanley, 2000; Mazzoni et al., 2002). Because flash floods determine high density-independent mortality of young fish (Schlosser, 1985; Magalhães et al., 2003), a continuous or extended reproductive period would increase the probability of successful recruitment (Matthews, 1998). Nevertheless, species other than C. pterostictum also display a clearly seasonal reproduction in variable environments (e.g., Alkins-Koo, 2000; Paugy, 2002; Mazzoni & Silva, 2006), so that the relation between life-history patterns and flash floods is unclear.
The results for C. pterostictum indicate the existence of small scale variability in life-history within the same stream. However, they did not support the hypothesis that fish in the most severely affected site should exhibit higher investment in present reproduction, with a trade-off for smaller body size and lower condition (see Table 3). This hypothesis derives from the general prediction that fish from environments that cause high density-independent mortality should exhibit shorter life-span, earlier maturity and higher reproductive investment (Schlosser, 1990; Danylchuk & Tonn 2006; Magalhães et al., 2003), which could be expected in PA, because of the more severe effects of flash floods. On the contrary, the results indicated a trade-off, in which less energy is allocated to reproduction (lower GSI) than to growth (larger size, larger size at maturity, and higher condition), while the opposite situation occurred in PB. In PA, besides being higher, condition did not change during the reproductive peak, suggesting that the degree of reproductive output (as expressed by the GSI) did not affect individual well-being. In PB, a higher reproductive cost was observed in females, which exhibited the lowest condition during the reproductive peak (same as observed for C. lauroi and C. alipioi by Braga, 2005).
The later maturity and lower instant reproductive investment observed in PA, can be interpreted as an alternative local change in life history. This change can be expected to occur in environments "prone to unpredictable, severe floods that kill eggs or larvae (Harvey, 1987)" (Matthews, 1998, p. 428), where higher probability of adult survivorship would be expected to result in additional cohorts in the future, then compensating for a present reduction in reproductive success. Higher investment in growth and condition may favor individual survival in PA, since larger individuals seem to be more resilient to flash floods (see Harvey, 1987), with GSI data suggesting a cost over reproduction. Higher condition would be advantageous because macroinvertebrate abundance usually decreases after flash floods, so that food availability would be diminished at least for a short period. In PA, three days after a flash flood in Summer 2000, no C. pterostictum was captured and all podostemacean macrophytes had been wiped off the substrate.
There are other studies showing that small-scale patterns are not always consistent with theory. For example, Spranza & Stanley (2000) found no trade-off between condition and reproduction in three small cyprinid species when comparing sites that differed in environmental variability. Possibly, it is not the overall life-history pattern of species that holds relation with environmental characteristics, but the intra-populational plasticity in life-history tactics within the small-scale environmental mosaic faced by populations. Such small-scale heterogeneity in streams was theoretically expressed in the patch dynamics concept (Pringle et al., 1988). The type and magnitude hydrological disturbances vary within a given stream system, and can be responsible for significant within-stream heterogeneity, influencing resident fish populations (Ross et al., 1985; Spranza & Stanley, 2000 and references therein). For example, higher downstream displacement and mortality has been observed for fish under 10 mm (Harvey, 1987), so that density-independent mortality can be determinant if there is overlap between spawning (or recruitment) and flash floods (as in C. pterostictum). For example, Lukas & Orth (1995) observed that the temporal pattern of streamflow fluctuation appears to be the most important abiotic factor determining nesting success or failure for smallmouth bass Micropterus dolomieu (Centrarchidae).
In summary, two distinct life-history strategies seem to occur in the studied population of C. pterostictum, each leading to population persistence in habitats that differ in severity of flash floods effects. One is the "bet-hedging" strategy observed in PA, in which there is a maximization of reproductive life-span by improving the survival probability of adults (longer lifespan, larger size and extended reproductive lifespan, but smaller instantaneous reproductive output) (Matthews, 1998). The other, observed in PB, is the optimization of present reproductive success (shorter life-span and shorter reproductive life span, but larger instantaneous reproductive output).
Although the life-history of C. pterostictum tends to be opportunistic (sensu Winemiller, 1989; Mazzoni et al., 2002), the analysis at a finer scale (within-population) indicated that local habitat characteristics may determine a more complex pattern. Spatial variability in the effects of hydrological disturbances might influence life-history patterns within a stream, particularly where disturbance by flash floods is typical. Thus, the present study provides evidence that fish occupying more severely disturbed areas of the stream are not necessarily at an energetic or reproductive disadvantage (see Spranza & Stanley, 2000), but only under environmental conditions that favor distinct patterns of energy allocation and population persistence, what is consistent with life-history theory (Matthews, 1998). Therefore, phenotypic plasticity in life-history could be expected to be common in stream fish populations, particularly in species that are widespread in a stream system with variable or mosaic-like habitat characteristics.
Acknowledgements
We thank C. B. Fialho and M. P. Albrecht for suggestions to an earlier version of the manuscript, L. C. Kucharski for advice on data analysis and to two anonymous referees, whose contribution improved the manuscript. This work was partially supported by CAPES, WWF/USAID and PIBIC-CNPq/UFRGS (to SC) and a CNPq research grant to SMH (304036/2007-2).
Literature Cited
Accepted August 2008
Published December 22, 2008
- Alkins-Koo, M. 2000. Reproductive timing of fishes in a tropical intermittent stream. Environmental Biology of Fishes, 57:49-66.
- Anderson, R. O. & R. M. Neumann. 1996. Length, weight, and associated structural indices. Pp. 447-482. In: B. R. Murphy & D. W. Willis (Eds). Fisheries techniques, 2nd edition. Bethesda, American Fisheries Society.
- Aranha, J. M., D. F. Takeuti & T. M. Yoshimura 1998. Habitat use and food partitioning of the fishes in a coastal stream of Atlantic Forest. Revista Brasileira de Biologia Tropical, 46:951-959.
- Azevedo, M. A., L. R. Malabarba & C. B. Fialho. 2000. Reproductive biology of the inseminating glandulocaudine Diapoma speculiferum Cope (Teleostei: Characidae). Copeia, 2000(4):983-989.
- Becker, F. G. 2002a. Distribuição e abundância de peixes de corredeiras e suas relações com características de hábitat local, bacia de drenagem e posição espacial em riachos de Mata Atlântica (bacia do rio Maquiné, RS, Brasil). Unpublished Ph.D. Dissertation, Universidade Federal de São Carlos, São Carlos, 201p.
- Becker, F. G. 2002b. Observations on the reproduction, sex ratio and size composition of Trachelyopterus lucenai (Teleostei: Auchenipteridae) in lake Guaíba, RS, Brazil. Biociências, Porto Alegre, 9(2):85-96.
- Becker, F. G., G. V. Irgang, H. Hasenack, F. S. Vilella & N. F. Verani. 2004. Land cover and conservation state of a region in the southern limit of the Atlantic Forest (river Maquiné basin, Rio Grande do Sul, Brazil). Brazilian Journal of Biology, 64(3B):569-582.
- Buckup, P. A. 2003. Family Crenuchidae (South American darters). Pp. 87-95. In: Reis, R. E., S. O. Kullander & C. J. Ferraris, Jr. (Orgs.). Check list of the freshwater fishes of South and Central America. Porto Alegre, Edipucrs, 729p.
- Bozzetti, M. & U. H. Schulz. 2004. An index of biotic integrity based on fish assemblages for subtropical streams in southern Brazil. Hydrobiologia, 529:133-144.
- Braga, F. M. S. 2005. Spatial distribution of characidiin fishes (Teleostei, Crenuchidae) in the Ribeirão Grande system, a tributary of Paraíba do Sul river basin, Brazil. Acta Scientiarum (Biological Sciences), 27(3):259-263.
- Danylchuk, A. J. & W. M. Tonn. 2006. Natural disturbance and life history: consequences of winterkill on fathead minnow in boreal lakes. Journal of Fish Biology, 68(3):681-694.
- Hartz, S. M., A. C. Peret & G. Barbieri. 1999. Reproduction of Gymnogeophagus lacustris, a cichlid endemic to southern Brazil. Ichthyological Explorations of Freshwaters, 10:247-253.
- Harvey, B. C. 1987. Susceptibility of young-of-the year fishes to downstream displacement by flooding. Transactions of the American Fisheries Society 116:851-855.
- Hasenack, H. & L. W. Ferraro. 1989. Considerações sobre o clima da região de Tramandaí - RS. Pesquisas (Porto Alegre), 22:53-70.
- Le Cren, E. D. 1951. The length-weight relation and seasonal cycle in gonad weight and condition in Perch (Perca fluviatilis). Journal of Animal Ecology, 20:201-219.
- Link, J. S. & J. Burnett.2001. The relationship between stomach contents and maturity state for major northwest Atlantic fishes: new paradigms? Journal of Fish Biology, 59(4):783-794.
- Lobón-Cerviá, J. & P. A. Rincón. 2004. Environmental determinants of recruitment and their influence on the population dynamics of stream-living brown trout Salmo trutta Oikos, 105(3):641-646.
- Lukas, J. A. & D. J. Orth. 1995. Factors affecting nesting success of smallmouth bass in a regulated Virginia stream. Transactions of the American Fisheries Society, 124:726-735.
- Machado, M. D., D. C. Heins & H. L. Bart. 2002. Microgeographical variation in ovum size of the blacktail shiner, Cyprinella venusta Girard, in relation to streamflow. Ecology of Freshwater Fish, 11(1):11-19.
- Magalhães, M. F., I. J. Schlosser & M. J. Collares-Pereira. 2003. The role of life history in the relationship between population dynamics and environmental variability in two Mediterranean stream fishes. Journal of Fish Biology, 63:300-317.
- Matthews, W. J. 1998. Patterns in freshwater fish ecology. New York, Chapman & Hall, 756p.
- Mazzoni, R. & A. P. F. Silva. 2006. Aspectos da história de vida de Bryconamericus microcephalus (Miranda Ribeiro) (Characiformes, Characidae) de um riacho costeiro da Mata Atlântica, Ilha Grande, RJ. Revista Brasileira de Zoologia, 23(1):228-233.
- Mazzoni, R. & J. Petito. 1999. Reproductive biology of a Tetragonopterinae (Osteichthyes, Characidae) of the Ubatiba fluvial system, Maricá, RJ. Brazilian Archives of Biology and Technology, 42:455-461.
- Mazzoni, R., E. P. Caramaschi & N. Fenerich-Verani. 2002. Reproductive biology of a Characidiinae (Osteichthyes, Characidae) from the Ubatiba River, Maricá, RJ. Brazilian Journal of Biology, 62:487-494.
- Olsen, E. M. & L. A. Vollestad. 2003. Microgeographical variation in brown trout reproductive traits: possible effects of biotic interactions. Oikos, 100(3):483-492.
- Olsen, E. M. & L. A. Vollestad. 2005. Small-scale spatial variation in age and size at maturity of stream-dwelling brown trout, Salmo trutta Ecology of Freshwater Fish, 14(2):202-208.
- Paugy, D. 2002. Reproductive strategies of fishes in a tropical temporary stream of the Upper Senegal basin: Baoule River in Mali. Aquatic Living Resources, 15(1):25-35.
- Poff, N. & J. Ward. 1990. Physical habitat template of lotic systems: Recovery in the context of historical pattern of spatiotemporal heterogeneity. Environmental Management, 14(5):629-645.
- Pringle C. M., R. J. Naiman, G. Bretschko, J. R. Karr, M. W. Oswood, J. R. Webster, R. L. Welcomme & M. J. Winterbourn. 1988. Patch dynamics in lotic systems: the stream as a mosaic. Journal of the North American Benthological Society, 7,503-524.
- Ross, S. T., W. J. Matthews & A. A. Echelle. 1985. Persistence of stream fish assemblages: effects of environmental change. American Naturalist, 126:24-40.
- Schlosser, I. J. 1985. Flow regime, juvenile abundance, and the assemblage structure of stream fishes. Ecology 66:1484-1490.
- Schlosser, I. J. 1990. Environmental variation, life history attributes, and community structure in stream fishes: implications for management and assessment. Environmental Management, 14:621-628.
- Spranza, J. J. & E. H. Stanley. 2000. Condition, growth, and reproductive styles of fishes exposed to different environmental regimes in a prairie drainage. Environmental Biology of Fishes, 59(1):99-109.
- Vazzoler, A. E. 1996. Biologia da reprodução de peixes teleósteos: teoria e prática. Maringá, Nupelia-Eduem, 169p.
- Vila-Gispert, A. & R. Moreno-Amich. 2002. Life-history Patterns of 25 species from European freshwater fish communities. Environmental Biology of Fishes, 65(4):387-400.
- Winemiller, K. O. 1989. Patterns of variation in life-history among South American fishes in seasonal environments. Oecologia, 81:225-241.
- Winemiller, K. O. 2004. Life-history strategies of fishes. Pp. 106-107. In: Ray, G. C. & J. McComrick-Ray. (Eds.) Coastal-marine conservation: science and policy. Oxford, Blackwell Science.
- Zar, J. H. 1999. Biostatistical Analysis. Upper Saddle River, Prentice Hall.
Publication Dates
-
Publication in this collection
22 Jan 2009 -
Date of issue
Dec 2008
History
-
Received
Aug 2008 -
Accepted
22 Dec 2008