ABSTRACT:
The objective of the present study was to evaluate the digestibility and the immunomodulatory effects of diets supplemented with lutein in dogs. Sixteen adult dogs were distributed in two groups (control and test) in a completely randomized design. Dogs were fed for 120 days with iso-nutritive diets, which only difference was the inclusion of 45g lutein/kg in the diet fed to the test group. Blood samples were collected in the beginning and at the end of the experimental period to evaluate total lymphocyte, lymphocyte proliferation index, and CD4+ and CD8+ lymphocyte counts. In order to evaluate apparent diet digestibility, dogs were housed in metabolic cages for ten days (five days of adaptation and five for faeces total collection). Coefficients of total tract apparent digestibility were not different between dogs in the control and the lutein-fed groups. Lutein dietary supplementation did not influence total lymphocyte or lymphocyte proliferation index, but resulted in higher CD4+ and CD8+ lymphocyte numbers. Lutein supplementation did not affect diet digestibility or lymphocyte proliferation index in dogs. However, it increases the concentrations of CD4+ and CD8+ T-lymphocyte subtypes.
Key words:
carotenoids; CD4+; CD8+; immunity
RESUMO:
O objetivo do presente estudo foi avaliar a digestibilidade e os efeitos imunomoduladores de dietas suplementadas com luteína em cães. Dezesseis cães adultos foram distribuídos em dois grupos (controle e teste), em delineamento inteiramente casualizado. Os cães foram alimentados durante 120 dias com dietas iso-nutritivas, diferindo apenasna inclusão de 45g de luteína/kg na dieta do grupo teste. As amostras de sangue foram colhidas no início e no final do período experimental para avaliar o total de linfócitos, o índice de proliferação de linfócitos e a contagem de linfócitos CD4 + e CD8 +. Para avaliar a digestibilidade aparente da dieta, os cães foram alojados em gaiolas metabólicas por dez dias (cinco dias de adaptação e cinco para coleta total de fezes). Os coeficientes de digestibilidade aparente não foram diferentes entre cães do grupo controle e os alimentados com luteína. A suplementação dietética com luteína não influenciou a contagem de linfócitos totais ou o índice de proliferação de linfócitos, mas resultou em número de linfócitos CD4 + e CD8 + mais elevadas. A suplementação de luteína não afeta a digestibilidade da dieta ou o índice de proliferação de linfócitos em cães. No entanto, aumenta as concentrações dos subtipos CD4 + e CD8 + de linfócitos T.
Palavras-chave:
carotenoides; CD4+; CD8+; imunidade
INTRODUCTION:
The use of feedstuffs with functional properties may bring important health benefits to dogs, such as better immunity and more adequate intestinal environment (KIM et al., 2000aKIM, H.W. et al. Dietary lutein stimulates immune response in the canine. Veterinary Immunology Immunopathology, v.74, p.315-327, 2000a. Available: <Available: http://aleksabokarev.narod.ru/foreignarticle2/12.pdf
>. Accessed: Apr. 27, 2014.
http://aleksabokarev.narod.ru/foreignart...
; SWANSON et al., 2002SWANSON, K. et al. Supplemental fructooligosaccharides and mannanoligosaccharides influence immune function, ileal and total tract nutrient digestibilities, microbial populations and concentrations of protein catabolites in large bowel of dogs. Journal of Nutrition , v.132, p.980-989, 2002. Available: <Available: http://jn.nutrition.org/content/132/5/980.long
>. Accessed: Apr. 26. 2014.
http://jn.nutrition.org/content/132/5/98...
). Carotenoids are considered natural pigments that provide beneficial health properties both for humans and animals, including enhancement of immune system and reduced risk of degenerative diseases (ALVES-RODRIGUES & SHAO, 2004ALVES-RODRIGUES, A.; SHAO, A. The science behind lutein. Toxicology Letters, v.150, p.57-83, 2004. Available from: https://www.researchgate.net/publication/8632483>. Accessed: Apr. 24, 2014. doi: 10.1016/j.toxlet.2003.10.031.
https://www.researchgate.net/publication...
). Among carotenoids of the xanthophylls group, lutein and zeaxanthin (non-provitamin A compounds) are the main substances (VALDUGA et al., 2009VALDUGA E. et al. Produção de carotenóides: microrganismos como fonte de pigmentos naturais. Quimica Nova, v.32, p. 2429-2436, 2009. Available: <Available: http://www.scielo.br/pdf/qn/v32n9/v32n9a36.pdf
>. Accessed: Apr. 26. 2014.
http://www.scielo.br/pdf/qn/v32n9/v32n9a...
).
Lutein is obtained mainly from the extract of Tagetes erecta . Its molecular formula is C40H56O2, with fusion point at 190°C, and it is crystallized in the presence of ether and methanol (LOURENÇO, 2005LOURENÇO, M.L.G. Efeito da idade e da suplementação com luteína no hemograma, nas enzimas hepáticas, na glicemia e no proteinograma de neonatos felinos. 2005, p.193. Tese (Doutorado em Veterinária) - Universidade Estadual Paulista, Faculdade de Medicina Veterinária e Zootecnia, Botucatu, SP. Available: <Available: http://acervodigital.unesp.br/handle/unesp/163538?mode=full
>. Accessed: Apr. 26. 2014.
http://acervodigital.unesp.br/handle/une...
). Among the main benefits related to lutein, its antioxidant properties and immunomodulatory characteristics are highlighted, particularly its actions on blood T lymphocytes. Studies showed that dogs are able to absorb dietary lutein, which is almost entirely taken up by circulating lymphocytes, enhancing dogs' immune system and improving their general health status (STRINGHETA et al., 2006STRINGHETA, P.C. et al. Luteína: propriedades antioxidantes e benefícios à saúde. Alimentação e Nutrição, v.17, p.229-238, 2006. Available: <Available: https://www.researchgate.net/publication/49599824
>. Accessed: Apr. 26. 2014.
https://www.researchgate.net/publication...
). Lutein stimulated cell-mediated and humoral immune responses in dogs by increasing lymphocyte proliferation, particularly of CD5+, CD4+ and CD8+ T cells after stimulation with an antigen (KIM et al., 2000aKIM, H.W. et al. Dietary lutein stimulates immune response in the canine. Veterinary Immunology Immunopathology, v.74, p.315-327, 2000a. Available: <Available: http://aleksabokarev.narod.ru/foreignarticle2/12.pdf
>. Accessed: Apr. 27, 2014.
http://aleksabokarev.narod.ru/foreignart...
).
Lutein may present a cytoprotective role on gastrointestinal mucosa, presenting antioxidant and free radical scavenging activity and anti-inflammatory and immunomodulatory effects (KRISHNASWAMYet al., 2010KRISHNASWAMY, R. et al. Lutein protects HT-29 cells against Deoxynivalenol-induced oxidative stress and apoptosis: Prevention of NF-κB nuclear localization and down regulation of NF-κB and Cyclo-Oxygenase - 2 expression. Free Radical Biology and Medicine, v.40, p.50-60, 2010. Available: <Available: http://www.ncbi.nlm.nih.gov/pubmed/20347963
>. Accessed: Apr. 28, 2014. doi: 10.1016/j.freeradbiomed.2010.03.016.
http://www.ncbi.nlm.nih.gov/pubmed/20347...
). Besides, lutein can also aid the absorption of lipids (AMAR et al., 2004AMAR, I. et al. Microstructure transition derived from solubilization of lutein and lutein esters in food microemulsions. Colloids Surface Biointerference, v.33, p.143-150, 2004. Available from: https://www.researchgate.net/publication/223433557>. Accessed: Jun. 16, 2015. doi: 10.1016/j.colsurfb.2003.08.009.
https://www.researchgate.net/publication...
), because of its microemulsion form. Thus, it may be hypothesized that dietary supply of lutein may influence diet digestibility. Considering the beneficial effects of lutein described and the few studies with dogs, this study aimed to evaluate the diet digestibility and immunomodulation of dogs fed with diets supplemented with lutein.
MATERIALS AND METHODS:
Animals
The experiment was approved by the Ethical Committee for Animal Use of Agrarian Sciences of the Universidade Federal do Paraná, Curitiba, PR, Brasil. Sixteen healthy adult Beagle dogs of approximately 2 years of age and with 9.2±1.2kg (average ± standard deviation) body weight were fed for 120 days with two iso-nutritive diets, which only difference was the inclusion of 45g lutein kg-1 in the test diet. This inclusion level was recommended by the fabricant and the lutein source was from Marigold flower (Tagetes erecta ). The product contains 75% lutein and 2% zeaxanthin.
Digestibility
Dogs were divided into two groups of eight dogs, each group consisted of four males and four females. Dogs were fed with dry extruded experimental diets (Table 1): a control diet or a diet supplemented with 45g kg-1 active xanthophylls (lutein sourcewith 76.7% lutein and 5.2 zeaxanthin) extracted from marigold and rosemary, totalling eight replicates per treatment.
Diets were offered to dogs twice daily. The amount offered was calculated to supply metabolizable energy requirements (MER), estimated as according to the equation proposed by the NRC (2006NRC (NATIONAL RESEARCH COUNCIL). Nutrient Requirements of Dogs, Washington: National Academy of Sciences, 2006. 83p. (Report n° 8).) for adult dogs: MER (kcal day-1) = 130 x body weight0.75. Water was offered ad libitum.
Dogs were individually housed in 0.7m long x 0.6m high x 0.5m wide stainless steel metabolic cages for faeces collection. The experimental period consisted of five days of adaptation of dogs to the experimental diets, followed by five days of total faeces collection (AAFCO, 2004ASSOCIATION OF AMERICAN FEED CONTROL OFFICIALS (AAFCO). Dog and cat nutrient profiles. Oxford, 2004. (Official Publication of the Association of American Feed Control Officials Incorporated).). Dogs were weighed at the beginning and at the end of each period.
Faeces were collected twice daily, pooled per experimental treatment and stored in a freezer at -14oC. Feces were scored always by the same researcher according to a scale from 1 to 5 as follows: 1 = watery feces to 5 = hard, dry pellets (small, hard masses). A correction factor was applied to estimate energy loss in the urine, according to AAFCO (2004ASSOCIATION OF AMERICAN FEED CONTROL OFFICIALS (AAFCO). Dog and cat nutrient profiles. Oxford, 2004. (Official Publication of the Association of American Feed Control Officials Incorporated).). Pooled faecal samples were thawed to room temperature and individually homogenized. Faeces were dried in a forced-ventilation oven at 55oC (320-SE, Fanem, São Paulo, Brasil) until constant weight was achieved (72h), and then ground (Arthur H. Thomas Co., Philadelphia, PA, USA) to 1mm particle size.
Ground faecal samples were submitted to analyses to determine the coefficients of total tract apparent digestibility (CTTAD) of dry matter (DM), organic matter (OM) crude protein (CP, method 954.01), crude fibre (CF, method 962.10), ash (method 942.05), and ether extract in acid hydrolysis (EE, method 954.02) contents, according to the AOAC (1995). Dietary and faecal gross energy (GE) were determined using an isoperibol calorimeter (Parr Instrument Co., model 1261, Moline, IL, USA). All analyses were carried out in duplicate, with variation lower than 5% intra assay.
Blood analyses
Blood samples were collected in the beginning and at the end of the experiment, at the same hour, after dogs were fasted for 24 hours, by venipuncture of the external jugular vein using a 10ml syringe with a 25 x 7 needle.
The complete blood count (CBC) was determined using the routine techniques described by JAIN (1993JAIN, N.C. Essentials of veterinary hematology. Philadelphia: Lea & Febiger, 1993. 417p.). Total erythrocyte and leukocyte counts and haemoglobin concentration were determined using an automatic cell counter (Sysmex KX-21N, SYSMEX Corporation of America, Long Grove - USA).
Total plasma protein concentration was determined using a Goldberg manual refractometer. Differential leukocytes were counted in two slides per animal, which were prepared at the time of collection. Blood smears were stained with Wright stain and later an average of 200 cells was read per slide. At the end of the experimental period, additional blood samples were collected to analyse lymphocyte proliferation index and to count CD4+ and CD8+ lymphocytes. Peripheral blood mononuclear cells (PBMC) were obtained from a10mL aliquot of the collected blood stored in tubes containing sodium heparin. Blood aliquot was added in polypropylene tubes of 50ml, containing 10mL of Histopaque -1119. Tubes were centrifuged for 40min at 1410rpm (Eppendorf, Centrifuge 5810R, Hamburg - Germany) and then the layer of mononuclear cells between the Histopaque-1119 and plasma was removed and transferred to empty propylene tubes. The 20mL volume was completed with RPMI 1640 culture medium and 2% bovine faetal serum (BFS), and tubes were again centrifuged for 6min at 1200rpm. The supernatant was discarded and the same procedure was repeated twice to eliminate platelets. The remaining pellet was re-suspended in 2mL of culture medium [RPMI 1640, BFS at 10% SFB, antibiotics (100UmL-1 penicillin and 100mg mL-1 streptomycin), and glutamine 2mM]. Cell aliquot was placed in a CO2 oven at 37oC for at least 2 hours in culture tubes for stabilization in order to ensure cell viability. Cell viability was determined after 1:2 cell dilution in Tripan Blue stain (MCB Manufacturing Chemist Inc., Cincinnati, OH, EUA), after which cells were counted in a Neubauer chamber, with the aid of an optical microscope at 400 x magnification.
Lymphocyte proliferation index
After isolation, 106 cells mL-1 were cultivated in RPMI-1640 culture medium enriched with 10% BFS and 0.1% antibiotics (penicillin 10,000U mL-1 and streptomycin 10mg L-1) in 96-well plates (200mL final volume) at 37oC and an atmosphere of 95% air/5% CO2 for 48 hours. Lymphocytes were stimulated with 20mL solution of the mitogen Concanavalin A (Con A), which stimulates lymphocyte T proliferation. After 72h, 10µL of the reagent Vision Blue were added in each well, and the plate was again incubated for 2h. Fluorescence of each sample was read in a spectrofluorimeter.
The fluorophore was excited at 530nm and the emission was captured at 590nm. Lymphocyte proliferation index was obtained by dividing the values obtained in the wells with stimulation (ConA) by the values of the wells with no stimulation (RPMI medium).
Determination of CD4+ and CD8+ subtypes
CD4+ and CD8+ lymphocytes were counted by the method of flow cytometry. Rat anti-CD4 (DH29A) and anti-CD8 (CADO46A) monoclonal antibodies were used to quantify these cell populations in dogs. For immunofluorescence analysis, 5×105 cells were stored on culture plates maintained on ice. Cells were centrifuged at 1500rpm for 5min at 4oC, and the supernatant was discarded. Then, 10µL of the antibody solution was added to each sample, except negative controls. Cells were incubated for 30min in the dark at 4oC. A volume of 400µL of a binding buffer (2% BFS, 10mM Hepes, pH 7.4; 150mM NaCl; 5 mM KCl; 1mM MgCl2; 1.8mM CaCl2 , sterilized by filtration and stored at 4oC) were added. Cells were then read in a flow cytometer (FACS Calibur - BD biosciences, San Jose - CA - USA). Data were expressed as a percentage of positive cells corrected for the cells marked with the isotypical control antibody of flow cytometry.
Statistical analyses
Digestibility, proliferation of blood lymphocytes, and CD4+ and CD8+ data were analyzed according to a completely randomized design. The CBC data were analyzed according to a completely randomized design with a split plot arrangement, considering the effects of periods and diets. Each dog was considered one experimental unit, totaling eight replicate per treatment. All data were submitted to normality test (Shapiro-Wilk) to verify if their distribution was normal. Then the Student's t-test at P<0.05 significance level was appliedto digestibility, proliferation of blood lymphocytes, and CD4+ and CD8+ data. The CBC data were analyzed using the GLM procedures of SAS (SAS Inst. Inc., Cary, NC), considering the effects of periods and diets, at 5% significance level.
RESULTS:
Digestibility
No differences were observed in CTTAD or ME content between diets with or without lutein supplementation (P>0.05) (Table 2). All dogs presented similar intake of both diets (P>0.05).Body weight (mean = 9.2±1.2kg) and faecal score (mean = 3.4) were not influenced by the experimental treatments during the trial (P>0.05, data not shown).
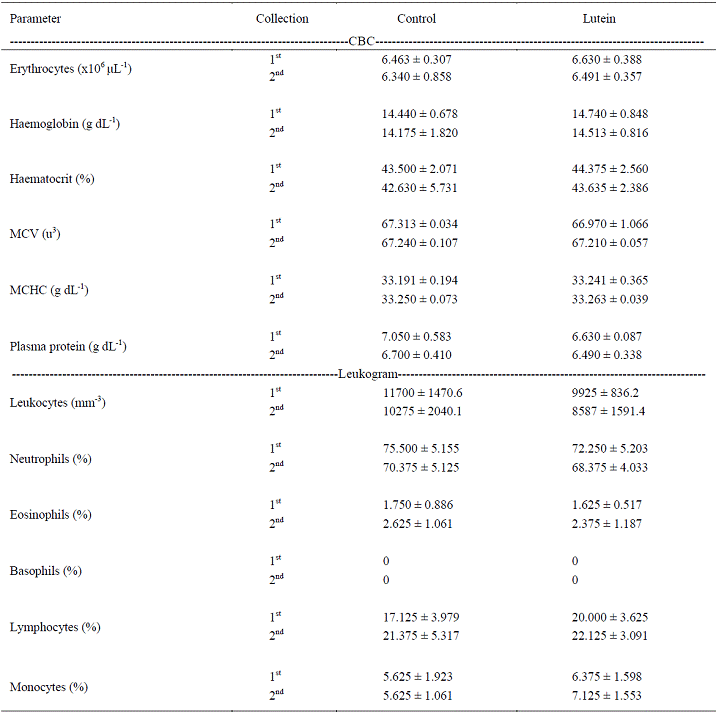
Blood parameters
No statistical differences (P>0.05) were verified between the control and the lutein group as to the evaluated blood count parameters (Table 3). Dietary lutein supplementation did not influence total blood counts (P>0.05). The observed values are within the reference range considered as normal for dogs, which demonstrated that dogs used in the present experiment were healthy.
Lymphocyte proliferation and CD4+ and CD8+ counts
There was no difference in lymphocyte proliferation between dogs fed with the control diet or the lutein-supplemented diet (P>0.05). As to CD4+ and CD8+ lymphocyte differential counts, the group fed with the diet supplemented with lutein presented greater counts of both lymphocyte subtypes as compared to the control group (P<0.05) (Table 4).
DISCUSSION:
No studies on the effect of dietary supplementation of lutein in dog foods digestibility were reported in literature to allow any comparison. However in humans, studies showed that lutein in the form of microemulsions (as in the present study) are efficient for absorption, and can also aid the absorption of other dietary components, such as lipids (AMAR et al., 2004AMAR, I. et al. Microstructure transition derived from solubilization of lutein and lutein esters in food microemulsions. Colloids Surface Biointerference, v.33, p.143-150, 2004. Available from: https://www.researchgate.net/publication/223433557>. Accessed: Jun. 16, 2015. doi: 10.1016/j.colsurfb.2003.08.009.
https://www.researchgate.net/publication...
). Lutein may also present cytoprotective role on gastrointestinal mucosa, presenting antioxidant and free radical scavenging activity and anti-inflammatory and immunomodulatory effects (KRISHNASWAMY et al., 2010KRISHNASWAMY, R. et al. Lutein protects HT-29 cells against Deoxynivalenol-induced oxidative stress and apoptosis: Prevention of NF-κB nuclear localization and down regulation of NF-κB and Cyclo-Oxygenase - 2 expression. Free Radical Biology and Medicine, v.40, p.50-60, 2010. Available: <Available: http://www.ncbi.nlm.nih.gov/pubmed/20347963
>. Accessed: Apr. 28, 2014. doi: 10.1016/j.freeradbiomed.2010.03.016.
http://www.ncbi.nlm.nih.gov/pubmed/20347...
), promoting gastrointestinal health. Despite of the beneficial effects related, we did not find differences on nutrient digestibility of the diet supplemented with lutein. In health animals fed with diets without pro antioxidants and/or pro inflammatory agents, these effects are not significant to influence diet digestibility.
In the present study no differences were reported on CBC in dogs fed with diets with 45g kg-1 active xanthophylls (34.51g lutein kg-1 of diet). KRUGER et al. (2002KRUGER, C.L. et al. An innovative approach to the determination of safety for a dietary ingredient derived from a new source: case study using a crystalline lutein product. Food Chemistry Toxicology, v.40, p.1535-1549, 2002. Available: <Available: http://www.ncbi.nlm.nih.gov/pubmed/12176081
>. Accessed: Apr. 26. 2014.
http://www.ncbi.nlm.nih.gov/pubmed/12176...
) also did not find any influence of lutein on CBC in rats after four weeks of diet supplementation with the great dose of 260mg kg-1 of diet. These results demonstrated that lutein did not influence CBC in animals, even in higher doses.
In studies on the action of carotenoids on the immune system, improved immune responses were obtained in rats, mice and lymphocyte cultures (JYONOUCHI et al., 1994JYONOUCHI, H. et al. Immunomodulating actions of carotenoids: enhancement of in vivo and in vitro antibody production to T- dependent antigens. Nutrition Cancer, v.21, p.47-58, 1994. Available: <Available: http://www.tandfonline.com/doi/abs/10.1080/01635589409514303
>. Accessed: Apr. 26, 2014. doi: 10.1080/01635589409514303.
http://www.tandfonline.com/doi/abs/10.10...
). BENDICH & SHAPIRO (1986BENDICH, A.; SHAPIRO, S.S. Effect of b-carotene and canthanxantin on the imunne response in rats. Journal of Nutrition, v.116, p.2254-2262, 1986. Available: <Available: http://www.ncbi.nlm.nih.gov/pubmed/3098935
>. Accessed: Jul. 23, 2013.
http://www.ncbi.nlm.nih.gov/pubmed/30989...
) reported that lutein obtained from targets suppressed the growth of mammary tumours and increased lymphocyte proliferation in rats. These authors also reported that the carotenoids astaxanthin, canxanthin, lutein, and ß-carotene stimulated animal immune response. Xanthophylls, such as lutein, are more potent immunostimulators than ß-carotene, and their primary action site in humoral immune response is similar to that of T-helper cells (KRAMER & BURRI, 1997KRAMER, T.R.; BURRI, B.J. Modulated mitogenic proliferative responsiveness of lymphocytes in whole-blood cultures after a low-carotene diet and mixed-carotenoid supplementation in women. American Journal of Clinical Nutrition, v.65, p.871-875, 1997. Available: <Available: http://ajcn.nutrition.org/content/65/3/871.abstract
>. Accessed: Apr. 28, 2014.
http://ajcn.nutrition.org/content/65/3/8...
). However, the physiological meaning of these observations is still not clear.
T-helper CD4+ is a humoral immune response indicator, which help other white blood cells in immunologic processes. The CD4+ interferes on stimulation and maturation of B lymphocytes and indicates how well the immune system is working. T-cytotoxic CD8+ is a trans membrane glycoprotein that acts as a co-receptor for the T cell receptor. It is predominantly expressed on the surface of cytotoxic T cells, so is an indicator for cellular immune response. These cells destroy virus-infected cells and tumors cells (VECCHIONE et al., 2002VECCHIONE, A. et al. Modulating immune responses with dendritic cells: an attainable goal in veterinary medicine? Vet Immunol Immunopathol, v.87, p.215-221, 2002. Available: <Available: http://www.sciencedirect.com/science/article/pii/S016524270200079X
>. Accessed: Apr. 27, 2014. doi: 10.1016/S0165-2427(02)00079-X.
http://www.sciencedirect.com/science/art...
, SHIRWAN et al., 2003SHIRWAN, H. et al. Chronic cardiac allograft rejection in a rat model disparate for one single class i mhc molecule is associated with indirect recognition by cd4+ t cells. Transplant Immunology, v.11, p.17-185, 2003. Available: <Available: http://europepmc.org/abstract/med/12799202
>. Accessed: Apr. 28, 2014. doi: 10.1016/S0966-3274(03)00004-2.
http://europepmc.org/abstract/med/127992...
).
The results obtained in the present experiment are consistent with those of KIM et al. (2000aKIM, H.W. et al. Dietary lutein stimulates immune response in the canine. Veterinary Immunology Immunopathology, v.74, p.315-327, 2000a. Available: <Available: http://aleksabokarev.narod.ru/foreignarticle2/12.pdf
>. Accessed: Apr. 27, 2014.
http://aleksabokarev.narod.ru/foreignart...
) as to the stimulation of the production ofCD4+ and CD8+ lymphocytes in dogs. Those authors studied different lutein dietary inclusion levels (0, 5, 10, and 20mg kg-1) and observed a significant increase in the counts of those lymphocyte subtypes after eight weeks of dietary supplementation of 5 to 20mg lutein. KIM et al. (2000aKIM, H.W. et al. Dietary lutein stimulates immune response in the canine. Veterinary Immunology Immunopathology, v.74, p.315-327, 2000a. Available: <Available: http://aleksabokarev.narod.ru/foreignarticle2/12.pdf
>. Accessed: Apr. 27, 2014.
http://aleksabokarev.narod.ru/foreignart...
) also reported a significant increase in total lymphocyte proliferation after five weeks of 5mg lutein inclusion in the diets of dogs, which; however, was not observed in the present experiment.
In another study, KIM et al. (2000bKIM, H.W. et al. Modulation of humoral and cell-mediated immune responses by dietary lutein in cats. Veterinay Immunology Immunopathology, v.73, p.331-341, 2000b. Available: <Available: https://www.researchgate.net/publication/223679869
>. Accessed: Apr. 27, 2014. doi: 10.1016/S0165-2427(00)00152-5.
https://www.researchgate.net/publication...
) evaluated the dietary supplementation of lutein in cats, and showed that in these animals, responses depend on both amount and time of supplementation. The authors observed an increase of both total lymphocyte proliferation and CD4+ and CD8+ production after four weeks of the highest level of lutein supplementation. Studying the effect of the supplementation of different β-carotene and lutein levels on the in-vitro proliferation of newly-hatched chick lymphocytes, HAQ et al. (1996HAQ, A. et al. Effect of β-Carotene, Canthaxanthin, Lutein, and Vitamin E on Neonatal Immunity of Chicks When Supplemented in the Broiler Breeder Diets. Avian Diseases, v.40, p.823-827, 1996. Available: <Available: http://ps.oxfordjournals.org/content/75/9/1092.short
>. Accessed: Apr. 26. 2014. doi: 10.3382/ps.0751092.
http://ps.oxfordjournals.org/content/75/...
) used tetrahydrofuran (THF) as a vehicle for the introduction of carotenoids in the culture medium. The authors concluded that β-carotene and lutein can only effectively increase in-vitro lymphocyte proliferation in amounts lower than 10-6 molar in the presence of THF.
HOSKINSON et al. (1990HOSKINSON, C.D. et al. Age-related changes in mitogen-induced lymphocyte proliferation and polymorphonuclear neutrophil function in the piglet. Journal of Animal Science, v.68, p.2471-2478, 1990. Available: <Available: https://www.animalsciencepublications.org/publications/jas/pdfs/68/8/2471
>. Accessed: Apr. 26. 2014.
https://www.animalsciencepublications.or...
) orally fed a mice with a lutein-rich marigold extract for four weeks and demonstrated an increase in lymphocyte proliferation after two weeks of supplementation. Lutein stimulated the proliferation of lymphocytes induced by mitogens, which action is not characteristic of β-carotene (KIM, 2000aKIM, H.W. et al. Dietary lutein stimulates immune response in the canine. Veterinary Immunology Immunopathology, v.74, p.315-327, 2000a. Available: <Available: http://aleksabokarev.narod.ru/foreignarticle2/12.pdf
>. Accessed: Apr. 27, 2014.
http://aleksabokarev.narod.ru/foreignart...
). In experiments with rats, the dietary supplementation of lutein reduced the growth of transplanted mammary tumours and increased lymphocyte proliferation (CHEW, 1995CHEW, B.P. Antioxidant vitamins affect food animal immunity and health. Journal of Nutrition , v.125, p.1804S-1808S, 1995. Available: <Available: http://jn.nutrition.org/content/125/6_Suppl/1804S.extract
>. Accessed: Apr. 14 , 2013.
http://jn.nutrition.org/content/125/6_Su...
; CHEW et al., 1996CHEW, B.P. et al. Effects of lutein from marigold extract on immunity and growth of mammary tumors in mice. Anticancer Research, v.16, p.3689-3694, 1996. Available: <Available: http://www.ncbi.nlm.nih.gov/pubmed/9042242
>. Accessed: Jun. 25, 2014.
http://www.ncbi.nlm.nih.gov/pubmed/90422...
; PARK et al., 1998PARK, J.S. et al. Dietary lutein absorption from marigold extract is rapid in Balb/c mice. Journal of Nutrition , v.128, p.1802-1806, 1998. Available: <Available: http://jn.nutrition.org/content/128/10/1802.full
>. Accessed: Apr. 26. 2014.
http://jn.nutrition.org/content/128/10/1...
). The lack of statistical improvement on lymphocyte proliferation with lutein supplementation in the present study may be due to the lack of diet and environment challenges and due to the experiment be conducted with health adult animals.
Despite the lack of statistical difference in lymphocyte proliferation values, the results on T lymphocytes obtained in the present study also indicated the beneficial immunomodulatory effect of lutein, improving cell-mediated immunity, suggesting its potential inclusion in dog foods to improve health.
CONCLUSION:
Dietary inclusion of lutein did not affect nutrients digestibility or CBC and lymphocyte proliferation indexes in dogs. However, it increases the concentrations of CD4+ and CD8+ T-lymphocyte subtypes, demonstrating the possible immunomodulatory effect of lutein. These are important findings that may improve the general health of dogs and deserves to be better studied, mainly with greater levels of dietary lutein supplementation and with animals with different health status (E.g.: infectious by virus and bacteria, and tumors), or in specifics life stages, as puppies and geriatric dogs.
BIOETHICS AND BIOSSECURITY COMMITTEE APPROVAL
Protocol was approved by Research Ethics Committee 051/2011.
REFERENCES:
- ALVES-RODRIGUES, A.; SHAO, A. The science behind lutein. Toxicology Letters, v.150, p.57-83, 2004. Available from: https://www.researchgate.net/publication/8632483>. Accessed: Apr. 24, 2014. doi: 10.1016/j.toxlet.2003.10.031.
» https://doi.org/10.1016/j.toxlet.2003.10.031.» https://www.researchgate.net/publication/8632483 - AMAR, I. et al. Microstructure transition derived from solubilization of lutein and lutein esters in food microemulsions. Colloids Surface Biointerference, v.33, p.143-150, 2004. Available from: https://www.researchgate.net/publication/223433557>. Accessed: Jun. 16, 2015. doi: 10.1016/j.colsurfb.2003.08.009.
» https://doi.org/10.1016/j.colsurfb.2003.08.009.» https://www.researchgate.net/publication/223433557 - ASSOCIATION OF AMERICAN FEED CONTROL OFFICIALS (AAFCO). Dog and cat nutrient profiles. Oxford, 2004. (Official Publication of the Association of American Feed Control Officials Incorporated).
- ASSOCIATION OF OFFICIAL ANALYTICAL CHEMISTS (AOAC). Official Methods of Analysis.16.ed. Washington, 1995. 1025 p.
- BENDICH, A.; SHAPIRO, S.S. Effect of b-carotene and canthanxantin on the imunne response in rats. Journal of Nutrition, v.116, p.2254-2262, 1986. Available: <Available: http://www.ncbi.nlm.nih.gov/pubmed/3098935 >. Accessed: Jul. 23, 2013.
» http://www.ncbi.nlm.nih.gov/pubmed/3098935 - CHEW, B.P. Antioxidant vitamins affect food animal immunity and health. Journal of Nutrition , v.125, p.1804S-1808S, 1995. Available: <Available: http://jn.nutrition.org/content/125/6_Suppl/1804S.extract >. Accessed: Apr. 14 , 2013.
» http://jn.nutrition.org/content/125/6_Suppl/1804S.extract - CHEW, B.P. et al. Effects of lutein from marigold extract on immunity and growth of mammary tumors in mice. Anticancer Research, v.16, p.3689-3694, 1996. Available: <Available: http://www.ncbi.nlm.nih.gov/pubmed/9042242 >. Accessed: Jun. 25, 2014.
» http://www.ncbi.nlm.nih.gov/pubmed/9042242 - HAQ, A. et al. Effect of β-Carotene, Canthaxanthin, Lutein, and Vitamin E on Neonatal Immunity of Chicks When Supplemented in the Broiler Breeder Diets. Avian Diseases, v.40, p.823-827, 1996. Available: <Available: http://ps.oxfordjournals.org/content/75/9/1092.short >. Accessed: Apr. 26. 2014. doi: 10.3382/ps.0751092.
» https://doi.org/10.3382/ps.0751092.» http://ps.oxfordjournals.org/content/75/9/1092.short - HOSKINSON, C.D. et al. Age-related changes in mitogen-induced lymphocyte proliferation and polymorphonuclear neutrophil function in the piglet. Journal of Animal Science, v.68, p.2471-2478, 1990. Available: <Available: https://www.animalsciencepublications.org/publications/jas/pdfs/68/8/2471 >. Accessed: Apr. 26. 2014.
» https://www.animalsciencepublications.org/publications/jas/pdfs/68/8/2471 - JAIN, N.C. Essentials of veterinary hematology. Philadelphia: Lea & Febiger, 1993. 417p.
- JYONOUCHI, H. et al. Immunomodulating actions of carotenoids: enhancement of in vivo and in vitro antibody production to T- dependent antigens. Nutrition Cancer, v.21, p.47-58, 1994. Available: <Available: http://www.tandfonline.com/doi/abs/10.1080/01635589409514303 >. Accessed: Apr. 26, 2014. doi: 10.1080/01635589409514303.
» https://doi.org/10.1080/01635589409514303.» http://www.tandfonline.com/doi/abs/10.1080/01635589409514303 - KIM, H.W. et al. Dietary lutein stimulates immune response in the canine. Veterinary Immunology Immunopathology, v.74, p.315-327, 2000a. Available: <Available: http://aleksabokarev.narod.ru/foreignarticle2/12.pdf >. Accessed: Apr. 27, 2014.
» http://aleksabokarev.narod.ru/foreignarticle2/12.pdf - KIM, H.W. et al. Modulation of humoral and cell-mediated immune responses by dietary lutein in cats. Veterinay Immunology Immunopathology, v.73, p.331-341, 2000b. Available: <Available: https://www.researchgate.net/publication/223679869 >. Accessed: Apr. 27, 2014. doi: 10.1016/S0165-2427(00)00152-5.
» https://doi.org/10.1016/S0165-2427(00)00152-5.» https://www.researchgate.net/publication/223679869 - KRAMER, T.R.; BURRI, B.J. Modulated mitogenic proliferative responsiveness of lymphocytes in whole-blood cultures after a low-carotene diet and mixed-carotenoid supplementation in women. American Journal of Clinical Nutrition, v.65, p.871-875, 1997. Available: <Available: http://ajcn.nutrition.org/content/65/3/871.abstract >. Accessed: Apr. 28, 2014.
» http://ajcn.nutrition.org/content/65/3/871.abstract - KRISHNASWAMY, R. et al. Lutein protects HT-29 cells against Deoxynivalenol-induced oxidative stress and apoptosis: Prevention of NF-κB nuclear localization and down regulation of NF-κB and Cyclo-Oxygenase - 2 expression. Free Radical Biology and Medicine, v.40, p.50-60, 2010. Available: <Available: http://www.ncbi.nlm.nih.gov/pubmed/20347963 >. Accessed: Apr. 28, 2014. doi: 10.1016/j.freeradbiomed.2010.03.016.
» https://doi.org/10.1016/j.freeradbiomed.2010.03.016.» http://www.ncbi.nlm.nih.gov/pubmed/20347963 - KRUGER, C.L. et al. An innovative approach to the determination of safety for a dietary ingredient derived from a new source: case study using a crystalline lutein product. Food Chemistry Toxicology, v.40, p.1535-1549, 2002. Available: <Available: http://www.ncbi.nlm.nih.gov/pubmed/12176081 >. Accessed: Apr. 26. 2014.
» http://www.ncbi.nlm.nih.gov/pubmed/12176081 - LOURENÇO, M.L.G. Efeito da idade e da suplementação com luteína no hemograma, nas enzimas hepáticas, na glicemia e no proteinograma de neonatos felinos. 2005, p.193. Tese (Doutorado em Veterinária) - Universidade Estadual Paulista, Faculdade de Medicina Veterinária e Zootecnia, Botucatu, SP. Available: <Available: http://acervodigital.unesp.br/handle/unesp/163538?mode=full >. Accessed: Apr. 26. 2014.
» http://acervodigital.unesp.br/handle/unesp/163538?mode=full - NRC (NATIONAL RESEARCH COUNCIL). Nutrient Requirements of Dogs, Washington: National Academy of Sciences, 2006. 83p. (Report n° 8).
- PARK, J.S. et al. Dietary lutein absorption from marigold extract is rapid in Balb/c mice. Journal of Nutrition , v.128, p.1802-1806, 1998. Available: <Available: http://jn.nutrition.org/content/128/10/1802.full >. Accessed: Apr. 26. 2014.
» http://jn.nutrition.org/content/128/10/1802.full - SHIRWAN, H. et al. Chronic cardiac allograft rejection in a rat model disparate for one single class i mhc molecule is associated with indirect recognition by cd4+ t cells. Transplant Immunology, v.11, p.17-185, 2003. Available: <Available: http://europepmc.org/abstract/med/12799202 >. Accessed: Apr. 28, 2014. doi: 10.1016/S0966-3274(03)00004-2.
» https://doi.org/10.1016/S0966-3274(03)00004-2.» http://europepmc.org/abstract/med/12799202 - STRINGHETA, P.C. et al. Luteína: propriedades antioxidantes e benefícios à saúde. Alimentação e Nutrição, v.17, p.229-238, 2006. Available: <Available: https://www.researchgate.net/publication/49599824 >. Accessed: Apr. 26. 2014.
» https://www.researchgate.net/publication/49599824 - SWANSON, K. et al. Supplemental fructooligosaccharides and mannanoligosaccharides influence immune function, ileal and total tract nutrient digestibilities, microbial populations and concentrations of protein catabolites in large bowel of dogs. Journal of Nutrition , v.132, p.980-989, 2002. Available: <Available: http://jn.nutrition.org/content/132/5/980.long >. Accessed: Apr. 26. 2014.
» http://jn.nutrition.org/content/132/5/980.long - VALDUGA E. et al. Produção de carotenóides: microrganismos como fonte de pigmentos naturais. Quimica Nova, v.32, p. 2429-2436, 2009. Available: <Available: http://www.scielo.br/pdf/qn/v32n9/v32n9a36.pdf >. Accessed: Apr. 26. 2014.
» http://www.scielo.br/pdf/qn/v32n9/v32n9a36.pdf - VECCHIONE, A. et al. Modulating immune responses with dendritic cells: an attainable goal in veterinary medicine? Vet Immunol Immunopathol, v.87, p.215-221, 2002. Available: <Available: http://www.sciencedirect.com/science/article/pii/S016524270200079X >. Accessed: Apr. 27, 2014. doi: 10.1016/S0165-2427(02)00079-X.
» https://doi.org/10.1016/S0165-2427(02)00079-X» http://www.sciencedirect.com/science/article/pii/S016524270200079X
-
1
CR-2015-1493.R2
Publication Dates
-
Publication in this collection
03 Oct 2016 -
Date of issue
Dec 2016
History
-
Received
03 Nov 2015 -
Accepted
16 June 2016 -
Reviewed
17 Sept 2016